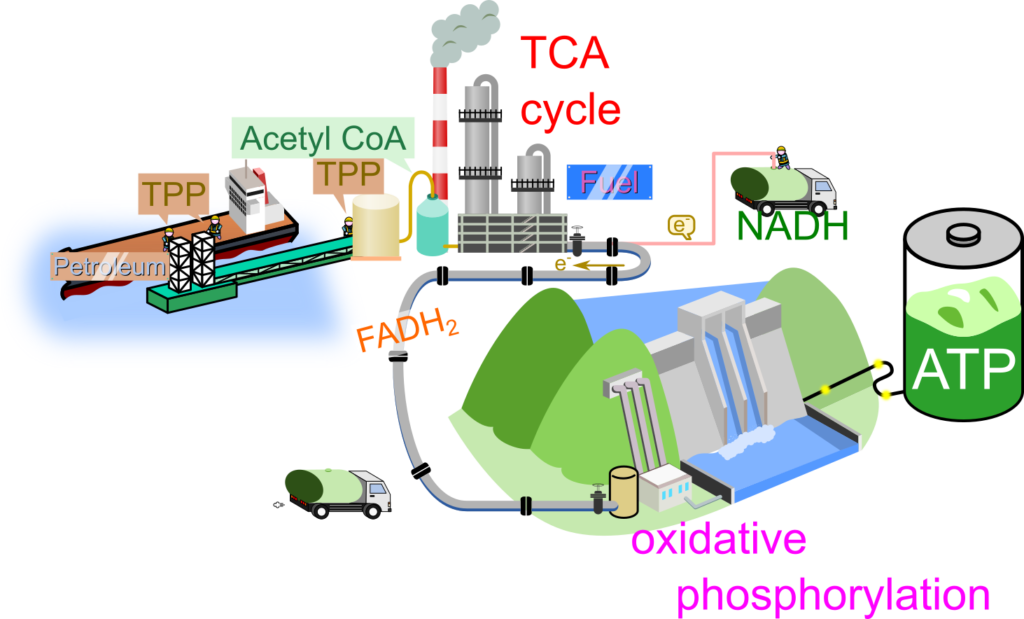
ビタミンB群には8種類が知られています。これらのビタミンの内、B1~B5はエネルギー代謝と深い関係を持っています。エネルギー代謝はTCA回路と酸化的リン酸化によって成り立っていますが、これらの経路は石油精製と揚水発電にたとえることができます。
解糖系によって生成されたピルビン酸は、チアミン二リン酸(TPP)とリポアミドの作用によって脱炭酸され、最終的にアセチル基として補酵素A(CoA)へ受け渡されます。ここで TPP はビタミンB1であるチアミンを含み、CoAはパントテン酸(ビタミンB5)を含みます。石油精製では、原油を前処理することで各成分を分離しやすい状態へ変換します。同様に、TPPはピルビン酸からアセチル基を取り出す反応に関与し、CoAはそのアセチル基を高反応性のチオエステルとして保持することで、続く反応を進行しやすくしています。
TCA 回路ではフラビン(ビタミンB2)を含むフラビンアデニンジヌクレオチドの還元型(FADH2)やナイアシン(ビタミンB3)を含むニコチンアミドアデニンジヌクレオチドの還元型(NADH)が生成されます。この過程は石油精製に近く、生成された FADH2 や NADH は石油精製で製造された燃料のように、電子伝達系に電子を伝達してエネルギーを供給します。
電子伝達系は揚水発電のポンプに相当し、伝達された電子を利用してミトコンドリアのマトリックス側から膜間腔へプロトンをくみ出して電荷勾配を作り出します。くみ出されたプロトンはくみ上げられた水のようにマトリックス側へ”落ちて”タービンを回すようにATP 合成酵素が ATP を生成するエネルギーを与えます。
このようにビタミン B1 ~ B5 はエネルギー代謝に深く関与している物質が多く含まれています。もちろんこれらのビタミンはエネルギー代謝だけでなくほかの代謝や異化経路でも活躍しています。ただし、これらビタミンの本質的な能力はエネルギー代謝でどう作用するのかを見ることで理解することができます。
この記事ではこれらのビタミン B1 ~ B5 について機能だけでなく、構造や吸収、異化経路について図解を中心にわかりやすくまとめています。
チアミン(ビタミンB1)
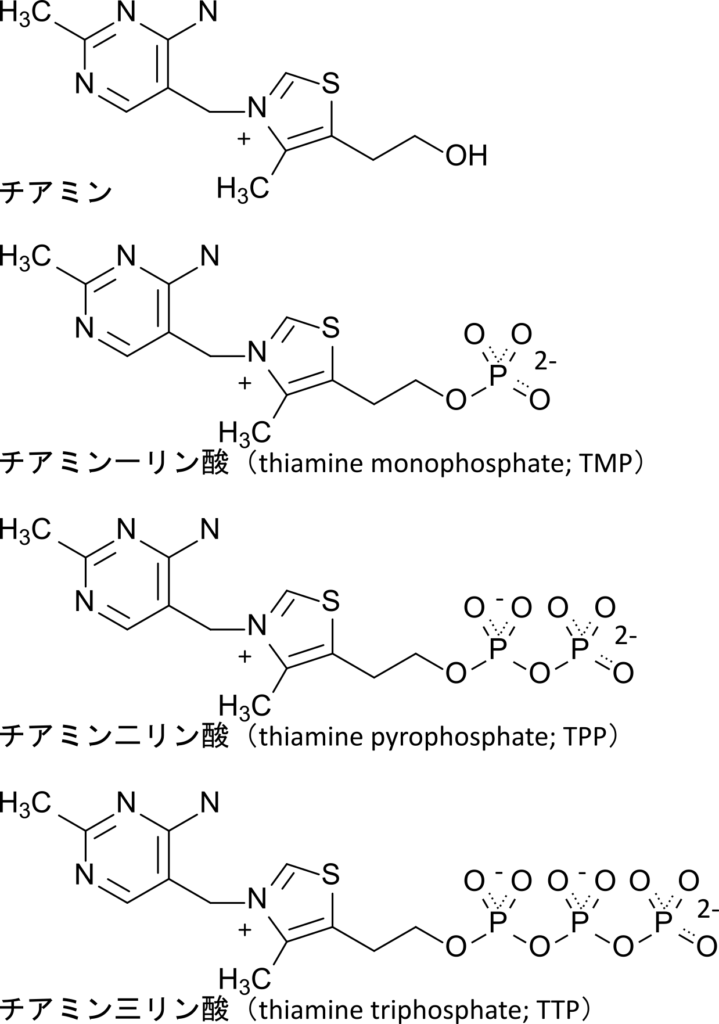
チアミンの誘導体であるチアミン二リン酸(TPP)はピルビン酸などの α-ケト酸の脱炭酸反応において補酵素として関与する物質です。この反応はTCA回路などのエネルギー代謝に重要です(図 1)。
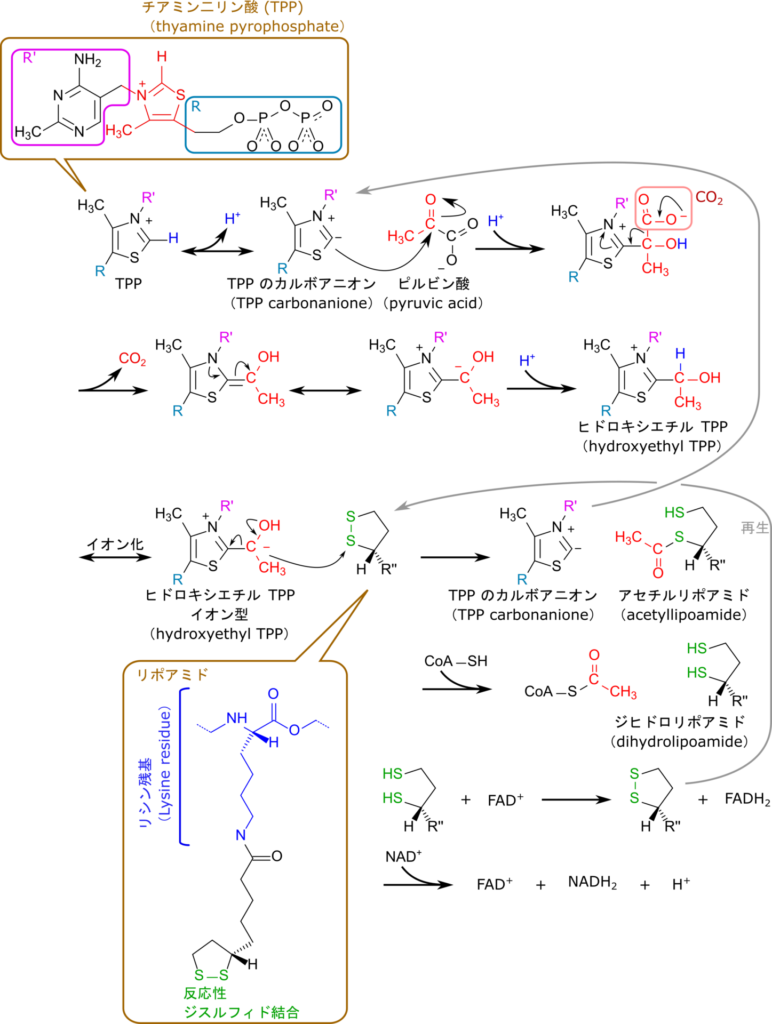
チアミンが関与する代表的な例としてピルビン酸の脱炭酸とCoA化を見ていきましょう。この反応はピルビン酸からCO2を脱離させた後、CoAとチオエステル結合を形成することでアセチルCoAを生成する反応で、アセチルCoAの供給に重要な反応です(図 2)。

この反応はチアミン二リン酸(TPP)からプロトンが脱離し、TPPのカルボアニオンが生成されることで始まります。カルボアニオンはピルビン酸のカルボニル基を攻撃して結合し、カルボキシル基が CO2 として脱離します。その後、電子の再配置を経てヒドロキシエチルTPPとなりますが、ヒドロキシエチル基(-CH(OH)-CH3)がリポアミドに転移されてアセチルリポアミドが生成されます。この時点でTPPは再生され、次の反応に利用可能となります。さらに、CoA のチオール基がアセチル基のカルボニル基を攻撃して結合しアセチル CoA を生成してリポアミドはジヒドロリポアミド(還元型)となります。ジヒドロリポアミドは FAD の作用により酸化されて再生されます。なお、FADH2 は NAD+ によって酸化されて再生されます。このようにしてチアミン二リン酸は TCA 回路などに対するアセチル CoA の供給に関与しています(図 3)。
フラビン(ビタミンB2)
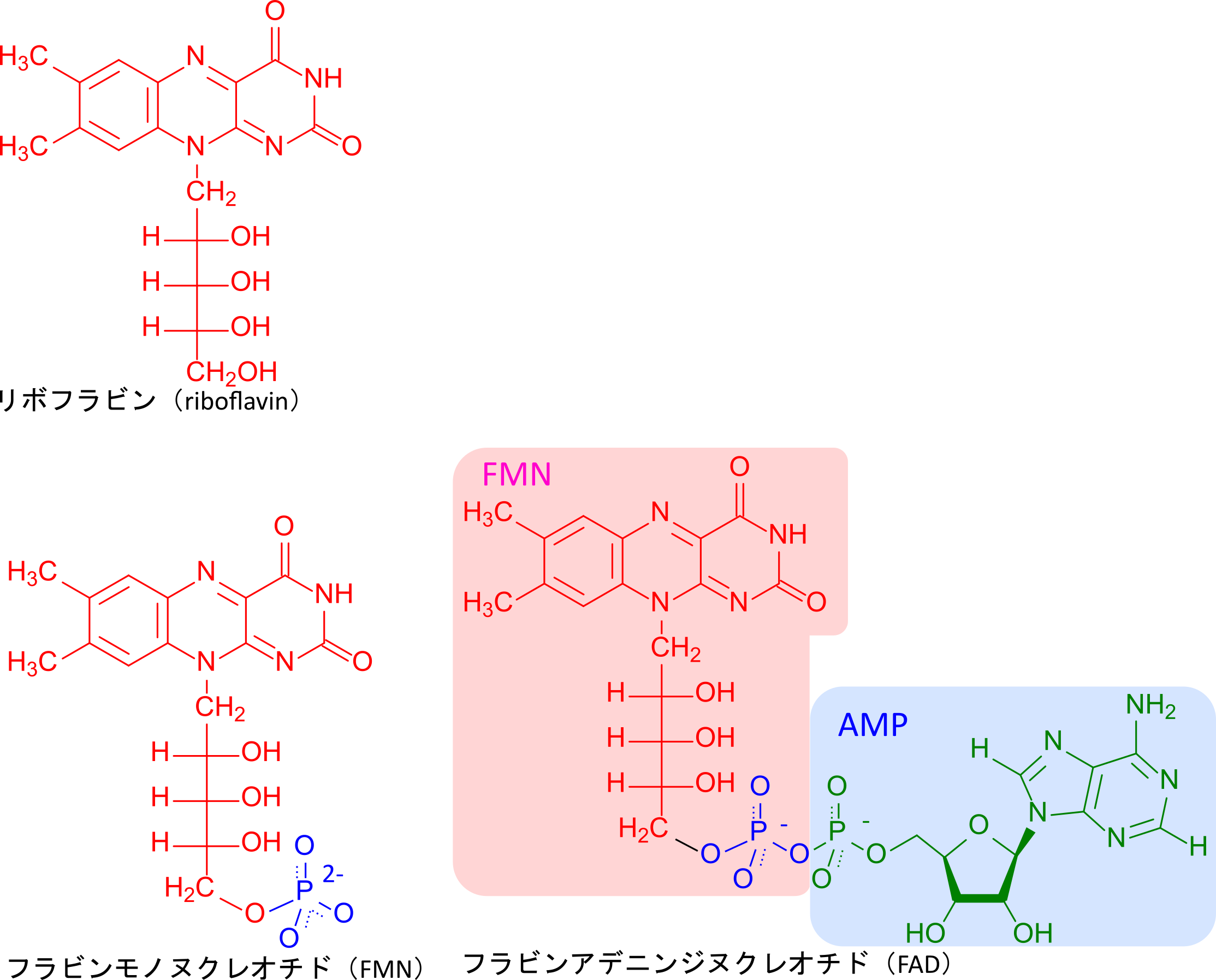
フラビンは生体内ではフラビンモノヌクレオチド(FMN)やフラビンアデニンジヌクレオチド(FAD)として存在し、TCA回路に関与するなどエネルギー代謝に重要な役割を担います(図 4)。これらの分子は酸化型と還元型を持っており、電子伝達などに関与しています(図 5)。
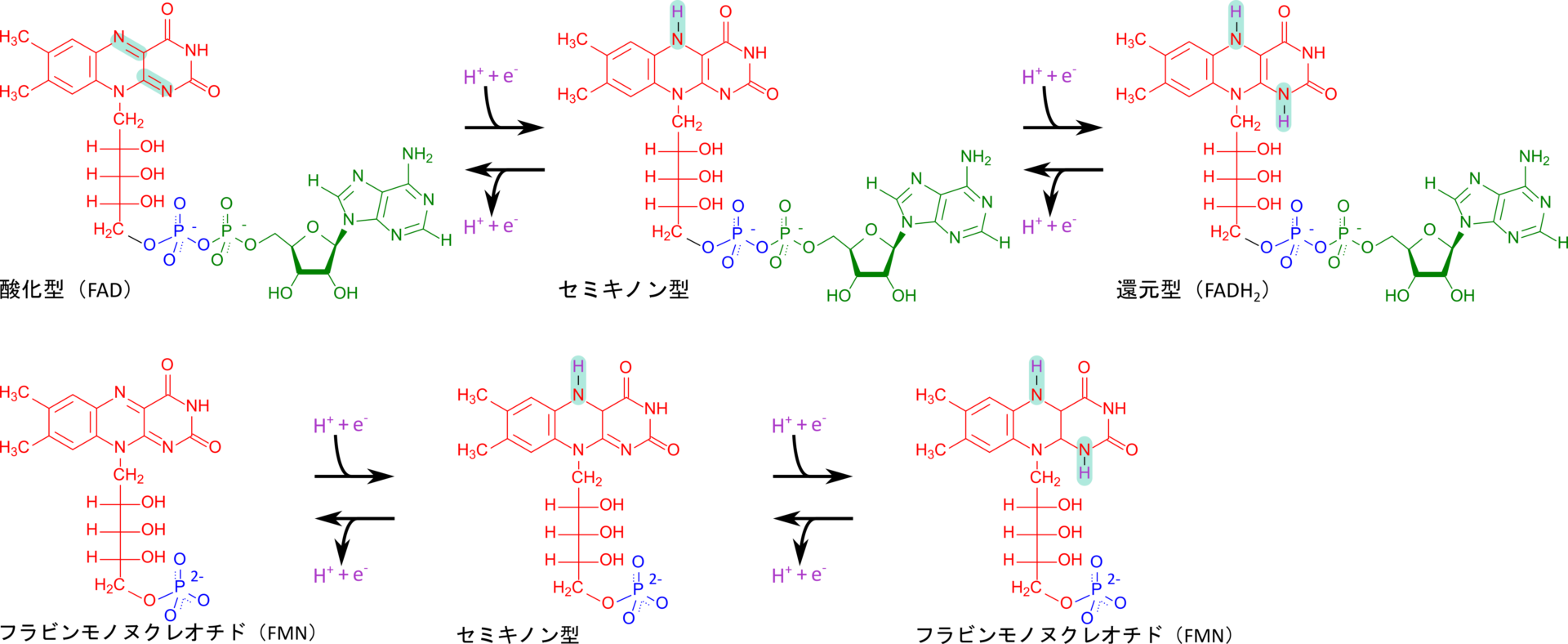
フラビンモノヌクレオチド(FMN)による電子のリレー
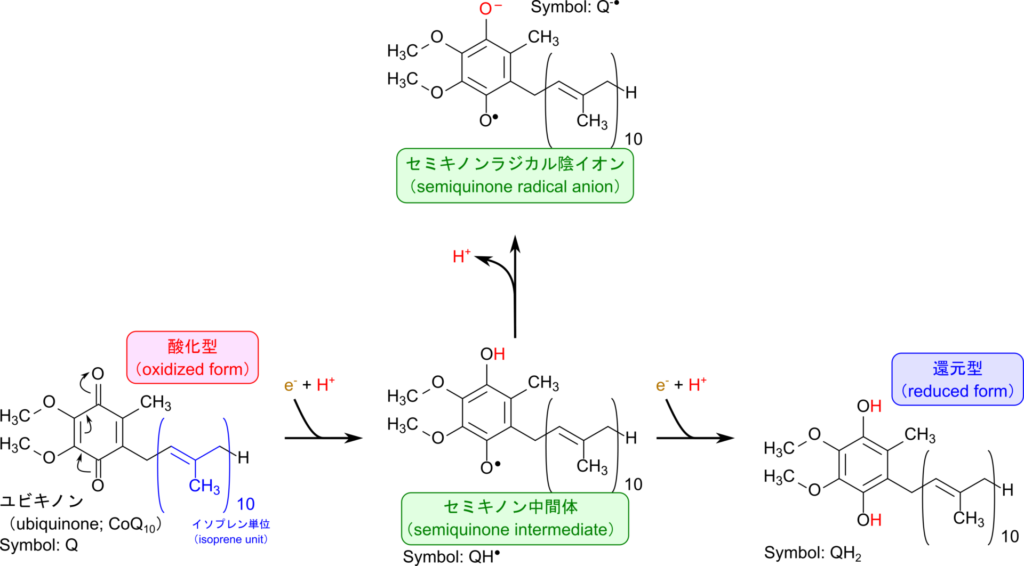
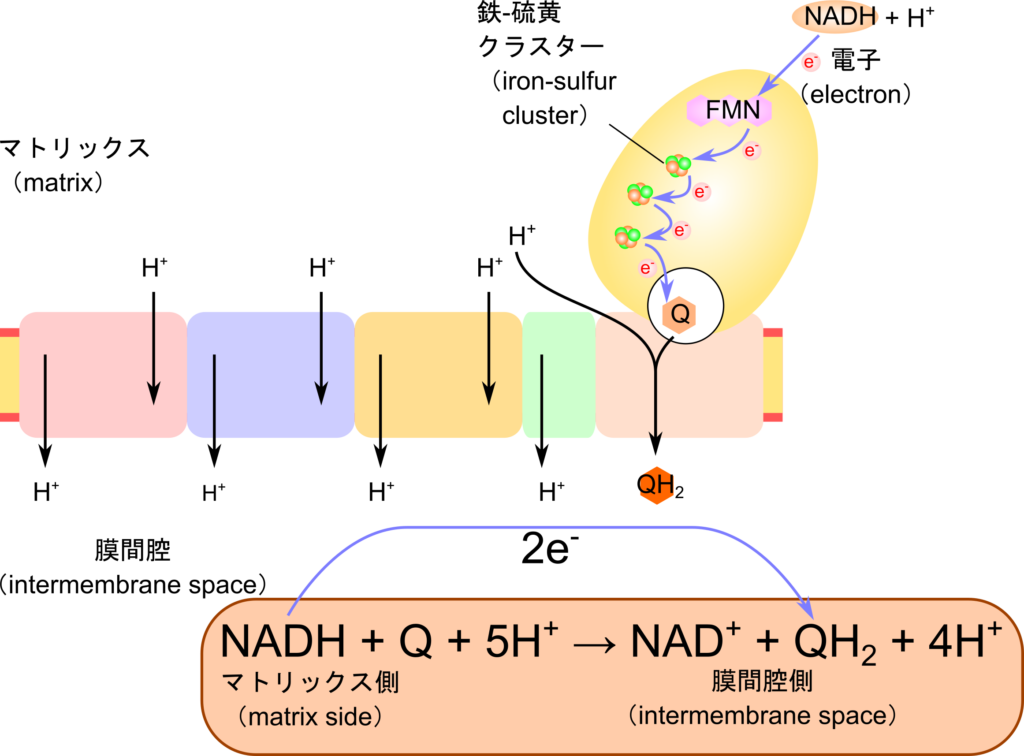
FMNは電子伝達系の複合体Ⅰの補欠分子族として存在し、NADHから受け取った電子を鉄-硫黄クラスターへ渡します。こうして伝達された電子は鉄-硫黄クラスターによりリレーされていき、ユビキノンに渡されます。電子を受け取ったユビキノンはセミキノン中間体を経て還元型ユビキノン(QH2)に変換されます(図 6)。この過程で還元されたユビキノンの負電荷によって複合体Ⅰは高次構造(立体構造など)変化や分子内のアミノ酸の pKa の変化などを引き起こし、ポンプのように複合体Ⅰがプロトンを汲み出すことに寄与します(図 7)。この過程はポンプに電気を流すことで駆動系が機能して水をくみ上げるようなイメージでとらえるとわかりやすいと思います。このようにして生み出されたプロトン濃度の差はATP 合成酵素がATPを生成するために利用されます。
このように FMN はTCA 回路などで産生されたNADHから電子を受け取って鉄-硫黄クラスターへ仲介する役割を担っています。
フラビンアデニンジヌクレオチド(FAD)による電子のリレー
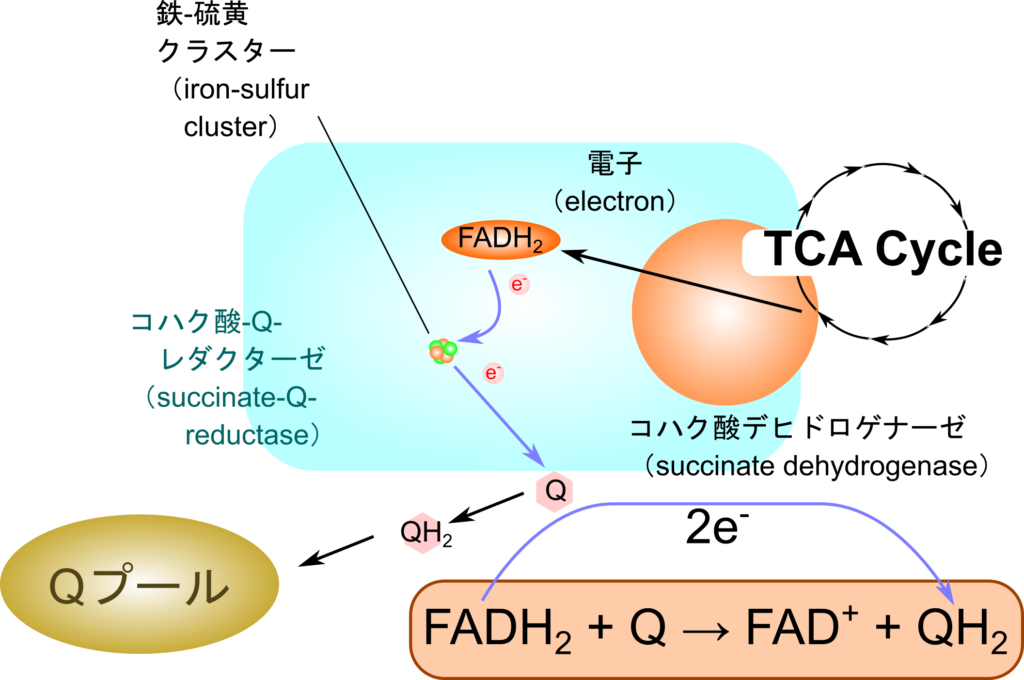
FAD は酸化型(FAD)と還元型(FADH2)が存在し、電子をリレーする際に利用されます。例えば、TCA 回路では、コハク酸が酸化されてフマル酸に変換される際に複合体 Ⅱ の作用により電子を電子伝達系に供給しますが、この経路で FAD が重要な役割を果たします。
FAD は複合体 Ⅱ のコハク酸デヒドロゲナーゼの補欠分子族として機能し、電子を受け取って、FADH2 に変換されます。FADH2 は電子伝達系の複合体 Ⅱ のFe-Sクラスターへと電子を渡し、自身は FAD へ再生されます(図 8)。こうして Fe-Sクラスターに渡された電子はユビキノン、複合体 Ⅲ、シトクロムc、複合体 Ⅳ へと順次渡されていき、最終的に酸素に渡されて複合体 Ⅲ と Ⅳ によるプロトンの汲み出しに寄与します。こうして生み出されたプロトン勾配を利用して ATP 合成酵素が ATP を産生します。なお、この経路は NADH が関与する経路とは異なり、複合体 Ⅰ を経由しないため、複合体 Ⅰ のプロトンのくみ出しには寄与しません。この違いは FADH2 あるいは NADH から生成できる ATP 分子数の差に影響しています。
ナイアシン(ビタミンB3)
ナイアシンは、ニコチン酸やニコチンアミドとして存在するビタミンであり、体内では NAD⁺ や NADP⁺ などの補酵素の構成成分として利用されます(図 9)。NAD+ や NADP+ には FAD と類似して酸化型と還元型が存在します。一方で、FAD が電子伝達を担うのに対してNAD+ や NADP+ は酸化型と還元型を相互変換することで酸化反応や還元反応を仲介します(図 10)。
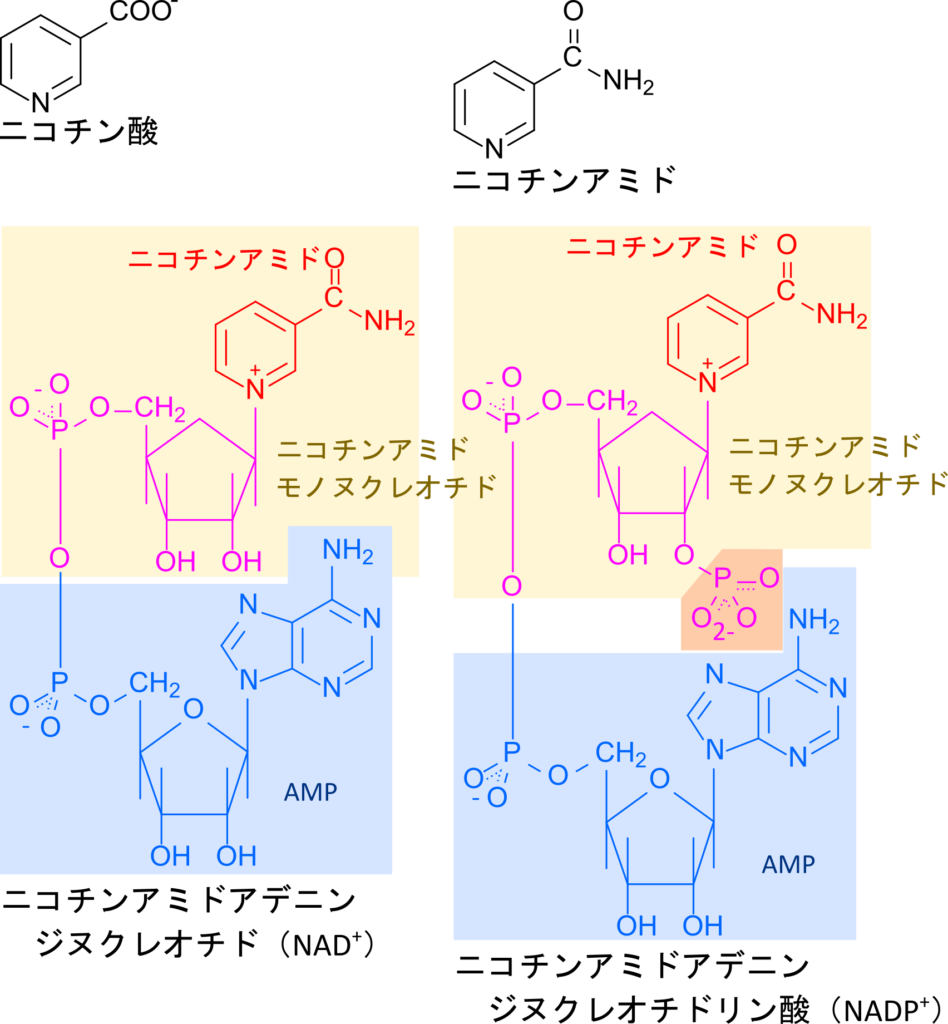
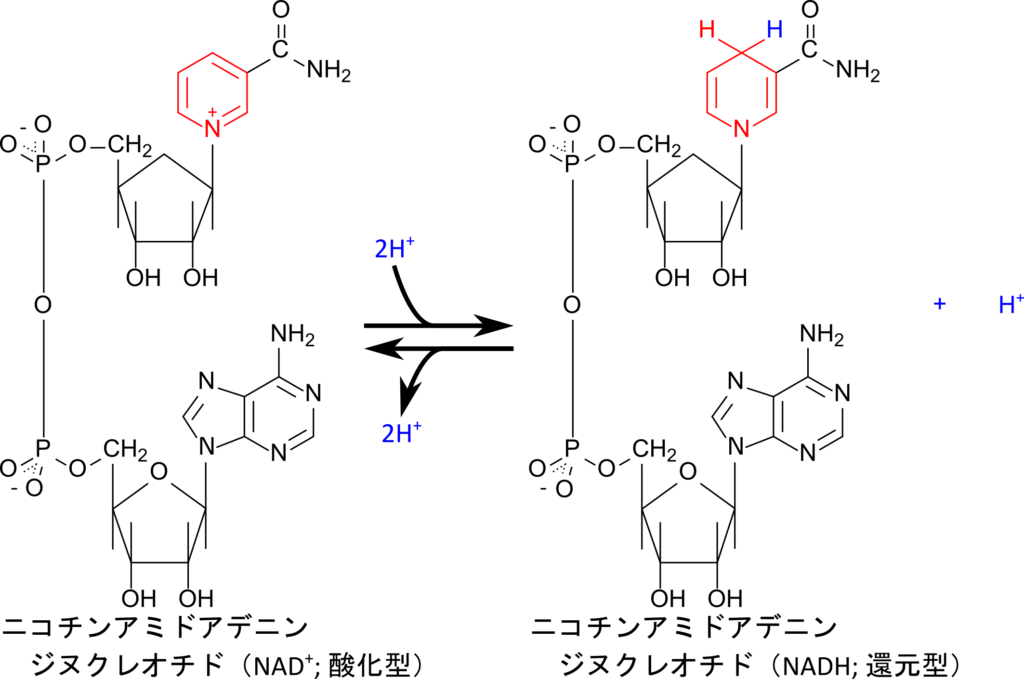
NAD+ 、NADP+ の機能と酸化・還元反応
NAD+ や NADP+ が関与する例を図 11に示します。
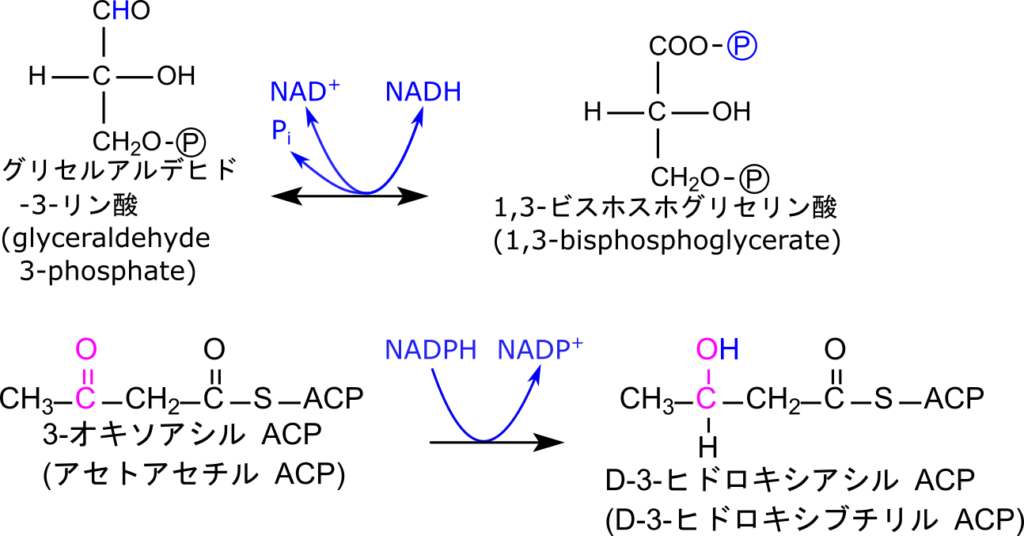
一つ目はグリセルアルデヒド-3-リン酸の酸化と共役したリン酸化です。この反応ではホルミル基を酸化してカルボキシル基に変換するとともにリン酸基を導入することで高エネルギー結合をもつ 1,3-ビスホスホグリセリン酸が生成されます。ホルミル基の酸化は NAD+ が関与して行われ、NADH が生成されます。さらに 1,3-ビスホスホグリセリン酸の高エネルギーリン酸基は ADP に転移させることで ATP を合成します。一方で、生成されたNADHは電子伝達系に電子を供給して酸化的リン酸化を通じて ATP 合成に関与するとともに NAD+ へ再生されます。このようにNAD+ は酸化反応を仲介すると同時に生成されたNADHがATP生成にも関与するなど、エネルギー代謝に重要な役割を果たします(図 11)。
もう一つの例は、脂肪酸合成における3-オキソアシルACPの還元反応です。この反応は3-オキソアシル ACPのケト基(C=O)を還元することで D-3-ヒドロキシアシルACP を生成します。このようにNADP+ は基質の還元反応に関与します(図 11)。
NAD+と NADP+ はともに酸化還元反応に関与する補酵素であり、その機能は非常に類似していますが、いくつか相違点もあります。NAD+ は解糖系やTCA回路などの代謝の酸化過程においてよく見られ、NADHは酸化的リン酸化において NAD+ に再生されます。一方で、NADPH は脂肪酸合成やステロイド合成などの還元反応を含む合成過程にみられます。反応に利用された NADPH はペントースリン酸回路を通じて再生されます。
NAD+ の代謝
NAD+ の合成経路
NAD+ の合成には以下の三つの経路が関与しています。
- de novo 経路
- Preiss-Handler 経路
- Salvage 経路
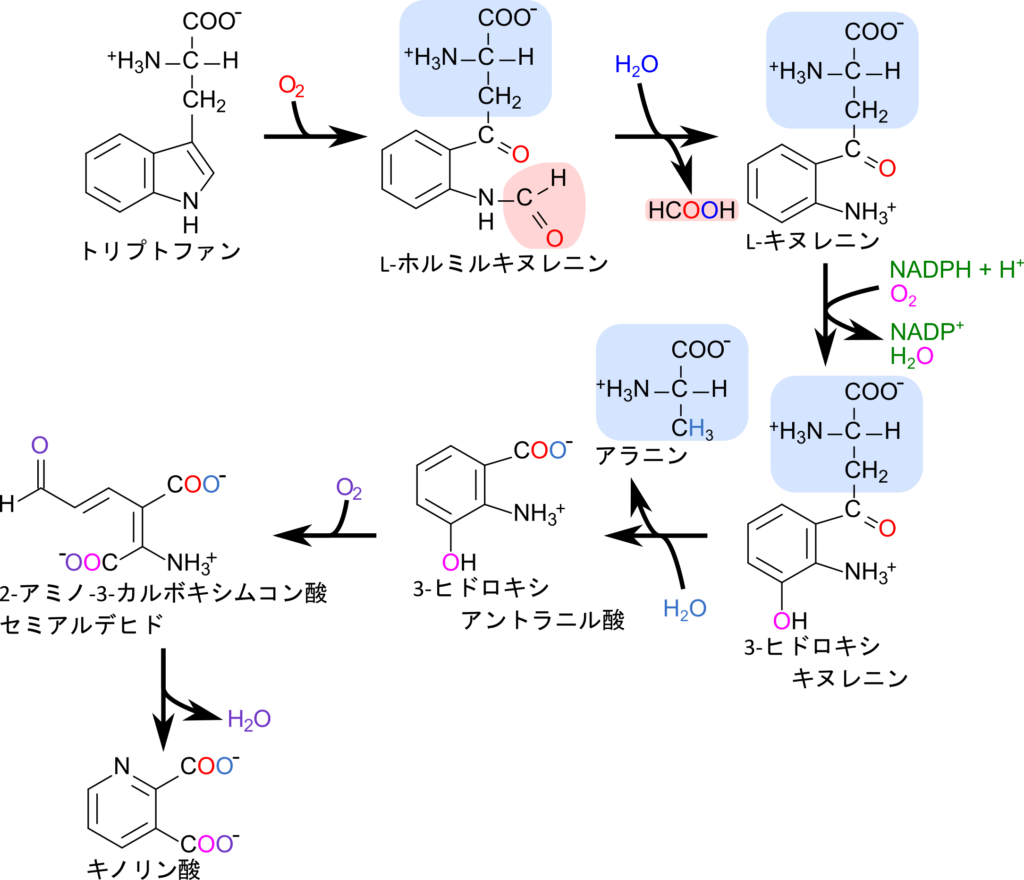
de novo 経路は新規にナイアシンを合成する経路です。トリプトファンから NAD+ が合成されます。トリプトファンからキヌレニンを経由してキノリン酸が生成されます(キヌレニン経路;kynurenine pathway)。キノリン酸に5-ホスホリボシル-1-二リン酸(5-Phosphoribosyl 1-diphosphate;PRPP)からホスホリボースが導入されるとニコチン酸-D-リボヌクレオチドが生成されます。さらにATP からAMP部分が導入され、グルタミンからアミノ基が転移されると NAD+ が生成されます(図 12、図 13)。
Preiss-Handler 経路は de novo 経路とよく似ています。de novo 経路ではキノリン酸とPRPPからニコチン酸-D-リボヌクレオチドが生成されて反応が進んでいきますが、Preiss-Handler 経路ではニコチン酸とPRPPからニコチン酸-D-リボヌクレオチドが生成されます(図 12、図 13)。
最後の経路は Salvage 経路です。Salvage 経路はニコチンアミドを再利用して NAD+ を合成する経路です。NAD+ はサーチュインやポリ ADP-リボースポリメラーゼ(PARP)、環状 ADP-リボース合成酵素(CD38やCD157)などに基質として利用されます。これらの酵素はヒストンなどの様々な細胞内タンパク質の脱アシル化やDNA修復、細胞内メッセンジャーとして機能する環状 ADP-リボース(cADPR)生成に関与します。この過程で生成されたニコチンアミドは PRPP からホスホリボースが、ATP から AMP 部分が供給されて NAD+ に再生されます(図 13)。
これら3つの経路ではde novo経路のみがナイアシンを新規に合成できます。Preiss-Handler 経路ではナイアシンの一つであるニコチン酸から合成が始まり、Salvage 経路ではニコチンアミドを出発物質としています。一方で de novo 経路では必須アミノ酸であるトリプトファンが出発物質として利用され、ナイアシンが合成されます。トリプトファン 60 mg から合成されるNAD+ 量は 1 mg のニコチンアミドから合成される量と同等です。このことからナイアシン当量という概念が考え出されました。ナイアシン当量は以下の通りです。![]()
ナイアシンはビタミンといいつつもトリプトファンから合成が可能です。しかし、トリプトファンは必須アミノ酸であり、トリプトファンの摂取量に余剰がある場合でもない限りナイアシンを摂取しなければ不足してしまいます。このことから通常ナイアシンはビタミンとみなされています。
ナイアシン由来化合物の排泄
NAD+ やニコチン酸、ニコチンアミドなどのナイアシン由来化合物は哺乳類ではN1-メチル-4-ピリドン-5-カルボキサミド(4PY)またはN1-メチル-2-ピリドン-5-カルボキサミド(2PY)として尿中に排泄されます。ナイアシン由来化合物は図 13のような経路をたどりニコチンアミドに変換されます。ニコチンアミドはS-アデノシルメチオニン(SAM)からメチル基を受け取って N1-メチルニコチンアミドに変換されます。その後、生成されたN1-メチルニコチンアミドが酸化を受けると4PYあるいは2PYに変換されます。これらの尿中における 2PY/4PY 比は種によって異なりますが、ヒトでは N1-メチル-2-ピリドン-5-カルボキサミドがナイアシン由来化合物の主要な排泄形態と考えられています。

パントテン酸(ビタミンB5)
パントテン酸は補酵素 A(Coenzyme A; CoA)やアシルキャリアタンパク質(Acyl Carrier Protein; ACP)の構成要素として知られています。パントテン酸はβアラニンとパントイン酸がアミド結合(-CO-NH-)したものです。さらに、パントテン酸のカルボキシル基とシステアミン(HS-CH2-CH2-NH2)がアミド結合し、一方で4’ 位(パントイン酸の4位)の水酸基がリン酸化を受けると 4’-ホスホパンテテインとなります。4’-ホスホパンテテイン酸がリン酸基を介して 5’-AMP と結合し、さらにリボースの 3’ 位にリン酸が結合すると CoA となります。一方で 4’-ホスホパンテテインのリン酸基を介してセリン残基の水酸基と結合すると ACP となります(図 15)。これらの分子は末端にチオール基(-SH)を有しており、アセチル基や脂肪酸のカルボキシル基を介して結合することができます。これらの分子は反応性の高いチオエステル(-CO-S-)を介して結合していることから、基質を活性化して反応しやすくさせたり、ACP のように基質を固定する役割を担っています。
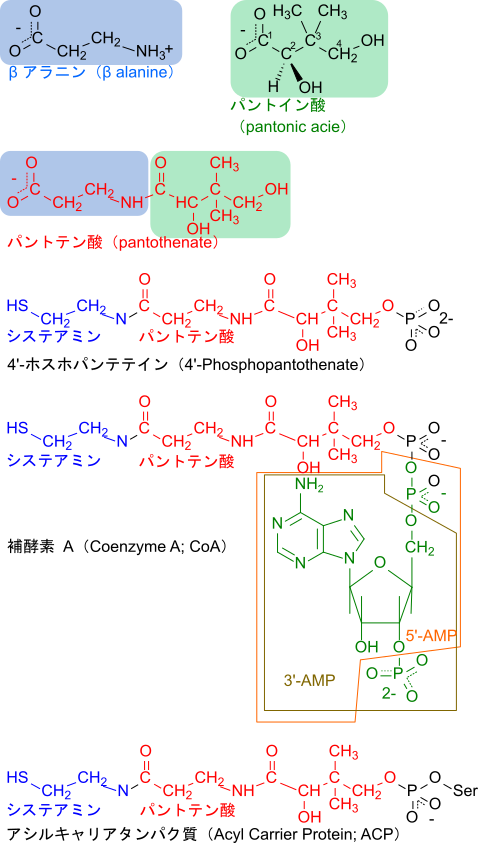
CoA やACP の生合成経路
CoA や ACP の合成ではまず 4’-ホスホパンテテインが合成されます。パントテン酸はパントテン酸キナーゼの作用により ATP を消費してリン酸化されると 4’-ホスホパントテン酸が生成されます。4’-ホスホパントテン酸はホスホパンテノイルシステインシンテターゼの作用によりシステインと縮合して4’-ホスホパンテノイルシステインが生成され、さらに ホスホパントテノイルシステインデカルボキシラーゼの作用により CO2 が脱離すると 4’-ホスホパンテテインとなります。システインとのカップリング反応では ATP が消費されて AMP となりピロリン酸を放出します。4’-ホスホパンテテインはデホスホ CoA ピロホスホリラーゼの作用により ATP から アデニリル基が転移されることでデホスホ CoA となります。最後にデホスホCoA キナーゼによりリボースの 3’ 位がATP を消費してリン酸化されると CoA が生成されます。生成された CoA の 4’-ホスホパンテテイン部分が ApoACP(4’-ホスホパンテテイン補欠分子族を持たないACP)のセリン残基の水酸基へ転移されて、3’, 5’-ADP が遊離することで ACP が生成されます。このようにして CoA や ACP が生成されます(図 16)。
CoA やACP の異化経路
ACP は ACP ホスホジエステラーゼの作用により加水分解を受けると 4’-ホスホパンテテインと ApoACP に分解されます。一方で CoA はエクトヌクレオチドピロホスファターゼ/ホスホジエステラーゼ(ENPP)などの作用により 3’, 5’-ADP を遊離させることで 4’-ホスホパンテテインに変換されます。4’-ホスホパンテテインが脱リン酸化を受けるとパンテテインとなり、パンテテインヒドロラーゼにより加水分解を受けてシステアミンが遊離してパントテン酸に異化されます(図 16)。
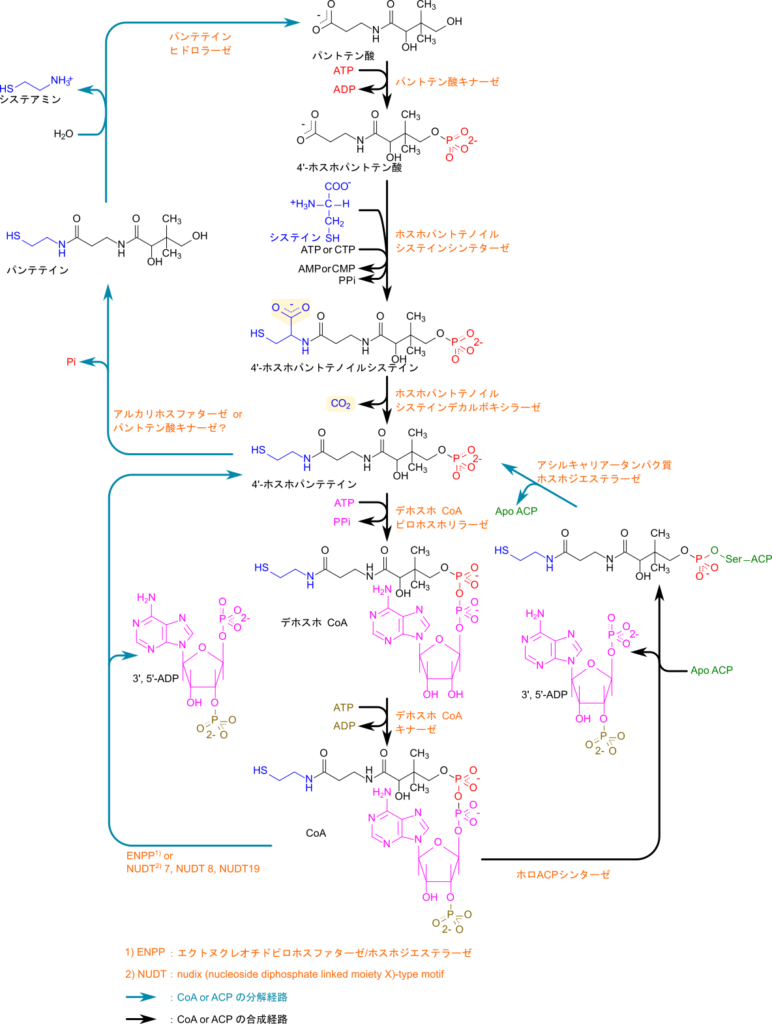
CoA や ACP の役割
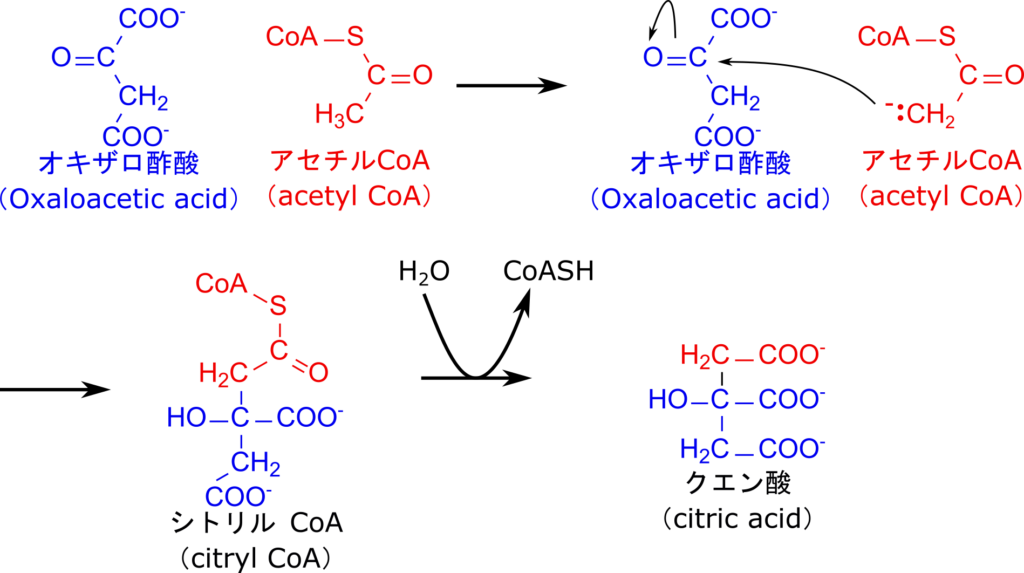
CoA や ACP は基質を活性化して反応性を高めたり、反応の場を与える役割を担います。例えば、TCA回路のアセチル CoA とオキサロ酢酸のカップリングでは、アセチル CoA の α プロトンが引き抜かれることでエノラートが生成され、オキサロ酢酸のカルボニル基を求核的に攻撃することで C-C 結合を形成してシトリル CoA を経て、最終的にクエン酸が生成されます(図 17)。この反応は、チオエステルのカルボニル基によって生成したエノラートが共鳴安定化されるため、α プロトンが引き抜かれやすくなることで進行します。
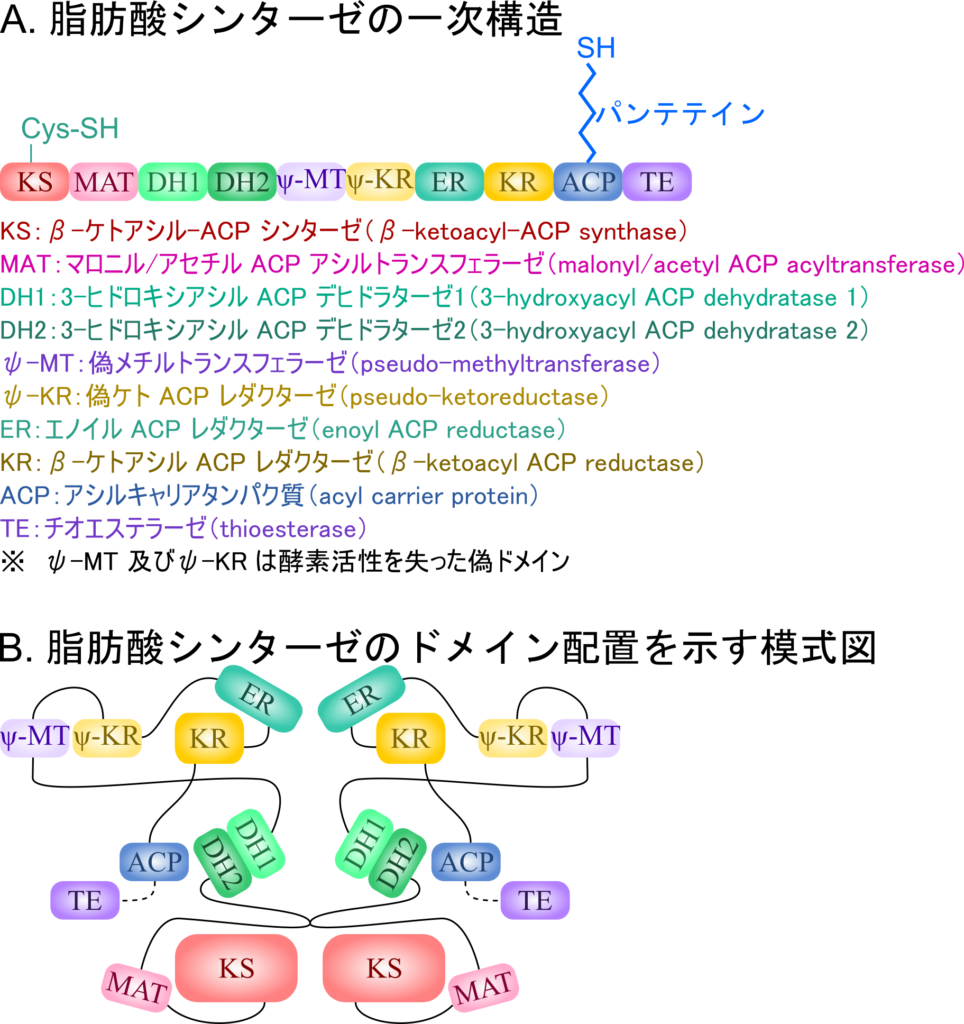
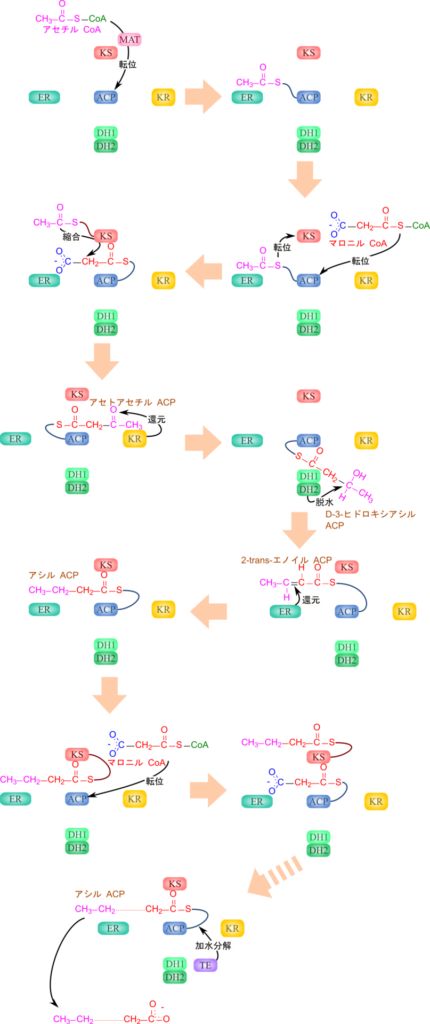
ACP はチオエステルを介してアセチル CoA のアセチル基を受け取ることで固定し、次いでアセチル 基を KS ドメインに移し、マロニル CoA からマロニル基を ACP が受け取ります。このようにすることでマロニル基とアセチル基を近接させて反応を促進します。さらに、ACP 上に中間体を保持することでこの後に続く還元反応や脱水反応などの反応を効率的に進行させる役割を担います。
このように CoA や ACP は、アシル基の反応性を高めるだけでなく、基質を酵素近傍に保持・運搬することで生体反応を効率化する役割を担っています(図 19)。
練習問題

ビタミンB群は、水溶性ビタミンとして知られるだけでなく、生体内では補酵素として機能し、エネルギー代謝を支える中心的な役割を担っています。チアミン二リン酸(TPP)による脱炭酸反応、FMNやFADによる電子伝達、NAD⁺やNADP⁺による酸化還元反応、CoAやACPによるアシル基転移反応など、ビタミンB群はATP産生や脂質・糖質代謝に深く関与しています。
一方で、これらのビタミンの役割はエネルギー代謝だけにとどまりません。DNA修復、脂肪酸合成、アミノ酸代謝、細胞内シグナル伝達など、多くの生命現象にも関与しています。生化学を学ぶうえでビタミンB群を理解することは、「代謝がどのように進行するのか」を理解することそのものとも言えます。
この記事では、ビタミンB群の種類や役割だけでなく、補酵素としての作用機序、代謝経路、生合成経路に至るまでを図解を用いて整理しました。エネルギー代謝や生化学を学ぶ際に、必要なときに何度でも読み返せる参考書として活用していただければ幸いです。
参考文献
共通
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 391-415, 459-484, 633-693
- D. Voet, J. G. Voet, C. W. Pratt 著、田宮信雄、八木達彦、遠藤斗志也、吉久徹訳 (2017) ボート 基礎生化学 第5版. 東京化学同人. pp. 294, 314-343, 477-513
- Peter J. Kennelly, Kathleen M. Botham, Owen P. McGuinness, Victor W. Rodwell, P. Anthony Weil 著、清水孝雄、水島昇 監訳 (2024). イラストレイテッド ハーパー・生化学 原書32版. 丸善出版. pp. 633-650
- 五十嵐脩、江指隆年 編 (2011). ビタミン・ミネラルの科学. 朝倉書店. pp. 1–39
ナイアシン
- Alemasova, E. E., & Lavrik, O. I. (2019). Poly(ADP-ribosyl)ation by PARP1: Reaction mechanism and regulatory proteins. In Nucleic Acids Research (Vol. 47, Issue 8, pp. 3811–3827). Oxford University Press. https://doi.org/10.1093/nar/gkz120
- Czura, A. W., & Czura, C. J. (2006). CD38 and CD157: Biological observations to clinical therapeutic targets. Molecular Medicine, 12(11–12), 309–311. https://doi.org/10.2119/2007-00006.Czura
- Mishra, K., & Kakhlon, O. (2025). The Crucial Role of NAD+ in MitochondrialMetabolic Regulation. In Biocell (Vol. 49, Issue 7, pp. 1101–1123). Tech Science Press. https://doi.org/10.32604/biocell.2025.061725
- North, B. J., & Verdin, E. (2004a). Protein family review Sirtuins: Sir2-related NAD-dependent protein deacetylases Gene organization and evolutionary history. http://genomebiology.com/2004/5/5/224
- Lenglet, A., Liabeuf, S., Bodeau, S., Louvet, L., Mary, A., Boullier, A., Lemaire-Hurtel, A. S., Jonet, A., Sonnet, P., Kamel, S., & Massy, Z. A. (2016). N-methyl-2-pyridone-5-carboxamide (2PY) — Major metabolite of nicotinamide: An update on an old uremic Toxin. In Toxins (Vol. 8, Issue 11). MDPI AG. https://doi.org/10.3390/toxins8110339