ビタミンCとビタミンEはいずれも抗酸化作用を発揮するビタミンであり、機能が類似しているだけでなく、互いに協調的に働く極めて関連性の深い分子です。細胞膜やLDLなどの脂質環境で起こる脂質酸化は火事のようなものであり、それ自体が脂質の劣化を招くだけでなく、火事が延焼するように周囲の脂質へと酸化反応を連鎖的に広げ、脂質酸化を増幅させてしまいます。
ビタミンEは脂溶性ビタミンであるため細胞膜などの脂質環境に局在し、脂質酸化の発端となる脂質ラジカルに働きかけて脂質酸化を抑制する、いわば消防士のような役割を担っています。一方でビタミンCは水溶性であるため液相で機能し、活性酸素種や脂質ラジカルなどのラジカル種を還元して生体を守る役割を担っています。ビタミンEが「消火」を担う消防士だとすれば、ビタミンCはトラブルメーカーとなるラジカル種を取り除く警察官のような役割だけでなく、酸化されたビタミンEを還元して再生する後方支援の役割も担っています。
このようにビタミンCとビタミンEは協調的に働くことで、細胞膜やリポタンパク質における脂質酸化を効率的に抑制しています。この記事では、これら抗酸化作用を有するビタミンについて、吸収・輸送、抗酸化作用のメカニズム、異化・排泄、さらには分解産物の機能まで含めて解説していきます。
ビタミンE
ビタミンEの構造と種類
ビタミンEには8種類が存在しています。これらの分子は大きく2種類に分類できて、一方がトコフェロール、もう一方がトコトリエノールです。これらの分子はクロマン環を有し、2位の炭素に炭素鎖が結合しています。トコフェロールではクロマン環に結合した炭素鎖は飽和炭素鎖であり、トコトリエノールは炭素鎖に三つの二重結合を持っています。クロマン環につくメチル基の数と位置によって以下のような α体、β体、γ体、δ体に分類されます(図 1)。
| 同族体 | R1 | R2 |
|---|---|---|
| α体 | メチル基(-CH3) | メチル基(-CH3) |
| β体 | メチル基(-CH3) | 水素(-H) |
| γ体 | 水素(-H) | メチル基(-CH3) |
| δ体 | 水素(-H) | 水素(-H) |

トコフェロールの生理活性ではα体がもっとも高く、次いでβ体、γ体、δ体の順に低くなります。α体の活性を 100 とすると相対的な生理活性の強さは以下のようになります。
- α体:100
- β体:40
- γ体:10
- δ体:1
この生理活性の強さは超低密度リポタンパク質(Very Low Density Lipoprotein;VLDL)中に取り込まれるときに必要とされる α-トコフェロール転送タンパク質(α-tocopherol transfer protein;α-TTP)との親和性によって決まります。α-TTPはα-トコフェロールと選択的に相互作用する分子です。そのため、α-トコフェロールが優先的にVLDLに取り込まれることになり、α-トコフェロールの末梢への輸送量が多くなり、α-トコフェロールの生理活性が大きくなります。つまり、ビタミンEの生理活性の強さはどれくらい末梢に輸送しやすいかに依存しています。
ビタミンEの吸収と輸送
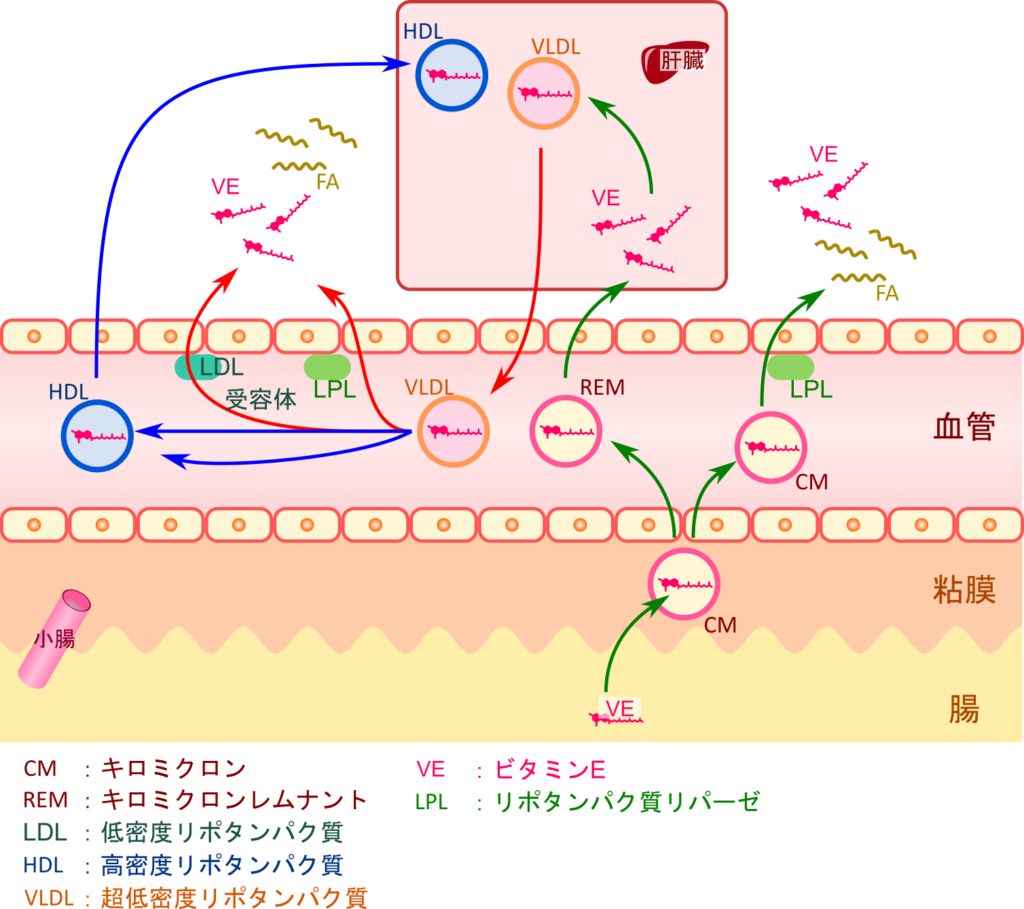
ビタミンEの取り込みと輸送は脂質と同様リポタンパク質を介して行われます。ビタミンEは粘膜中で他の脂質とともにキロミクロンに取り込まれて血中に放出されます。内皮細胞においてリポタンパク質リパーゼ(lipoprotein lipase;LPL)によりトリアシルグリセロールが分解されるとキロミクロンは小型化し、ビタミンEのごく一部が細胞内に移行します。一方で、大部分はキロミクロンレムナントとして肝臓に取り込まれます。肝臓に取り込まれたビタミン E はVLDL 中に導入されて、末梢細胞へ輸送される準備が整えられますが、α-TTPとの親和性によりVLDLに取り込まれるビタミンE量がα体、β体、γ体、δ体で異なります。内皮細胞のLPLはVLDL中のトリアシルグリセロールを分解して小型化することでIDL、LDLへ変換し、標的細胞による取り込みを誘導します。一方で、LDL受容体を介したエンドサイトーシスでもビタミンEが細胞内に取り込まれます。その後、ビタミンEはHDLに取り込まれて肝臓へ輸送されます(図 2)。
脂質酸化とビタミンEによる脂質酸化抑制
ビタミンEの最も重要な機能は抗酸化作用です。ビタミンEは脂溶性であるため細胞膜に局在することができ、脂質酸化を防止する役割を担います。
脂質酸化の発生メカニズム
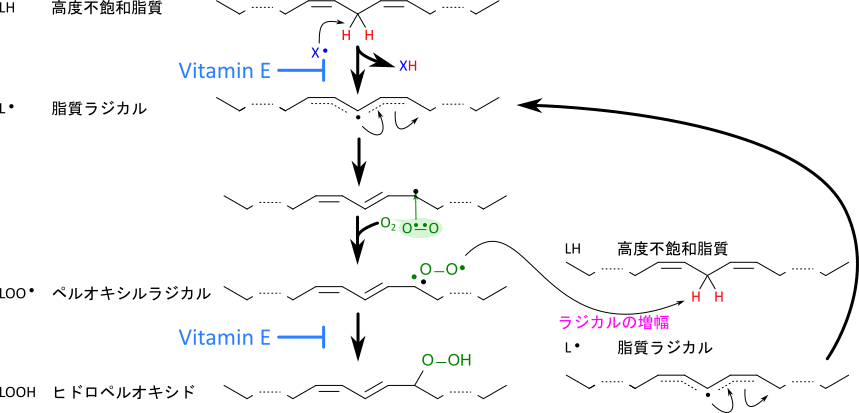
脂質酸化は不飽和脂肪酸がラジカル(X・)の攻撃を受けて脂質ラジカル(L・)を生成することから始まります。活性酸素などに代表されるラジカルは不対電子を持つため水素(H・)を抜き取り、脂質ラジカル(L・)を生成します。脂質ラジカルに酸素が付加するとペルオキシルラジカル(LOO・)を生成しますが、ペルオキシルラジカルはさらにほかの不飽和脂肪酸を攻撃して脂質ラジカル(L・)を生成します。このようにして脂質酸化は連鎖反応的に反応が進行してしまうため、反応が増幅されていきます。脂質酸化は最終的に脂質ヒドロペルオキシドを生成し、脂質の劣化を招いてしまいます(図 3)。
ビタミンEによる脂質酸化の抑制
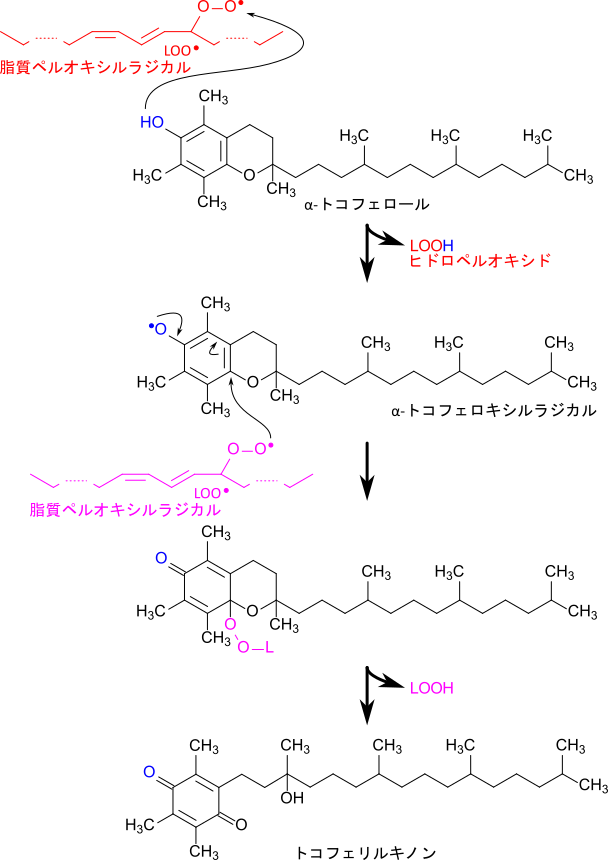
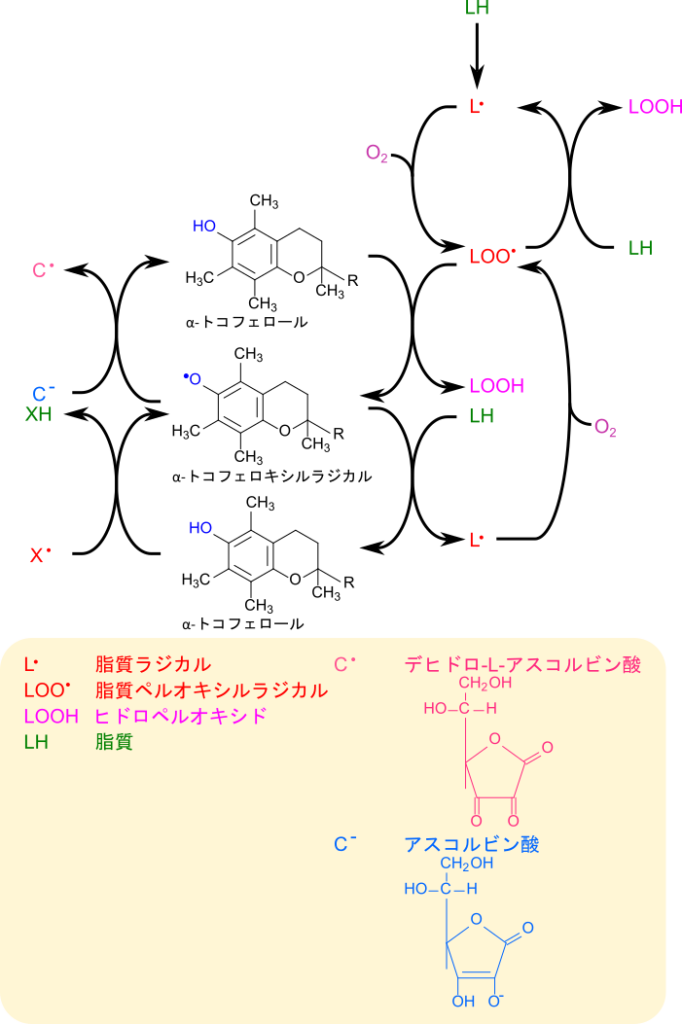
ビタミンEは活性酸素や脂質ペルオキシルラジカルなどのラジカルが水素を抜き取ることを阻害し、新たな脂質ラジカルが生成されることを防ぎます。図4に脂質ペルオキシルラジカルとα-トコフェロールの反応を示します。この反応は脂質ペルオキシルラジカルがトコフェロールのクロマン環の水酸基から水素を抜き取ることで始まります。このとき、脂質ペルオキシルラジカルは還元されてヒドロペルオキシドとなり、連鎖的なラジカル連鎖反応はいったん停止します。一方で、水素を抜き取られたα-トコフェロキシラジカルは他の脂質ペルオキシルラジカルなどのラジカル種との会合・中間体生成やさらなる酸化を経て、脂質を還元して脂質ヒドロペルオキシドに変換し、自身は酸化されトコフェリルキノンとなります(図 4)。あるいは、ビタミンCの作用により還元され、α-トコフェロールに再生されます。このようにビタミンEはラジカルに働きかけて、脂質酸化を早期に抑制することで脂質が劣化することを防いでいます。
ビタミンEの異化と排泄
ビタミンEの異化産物の種類と機能
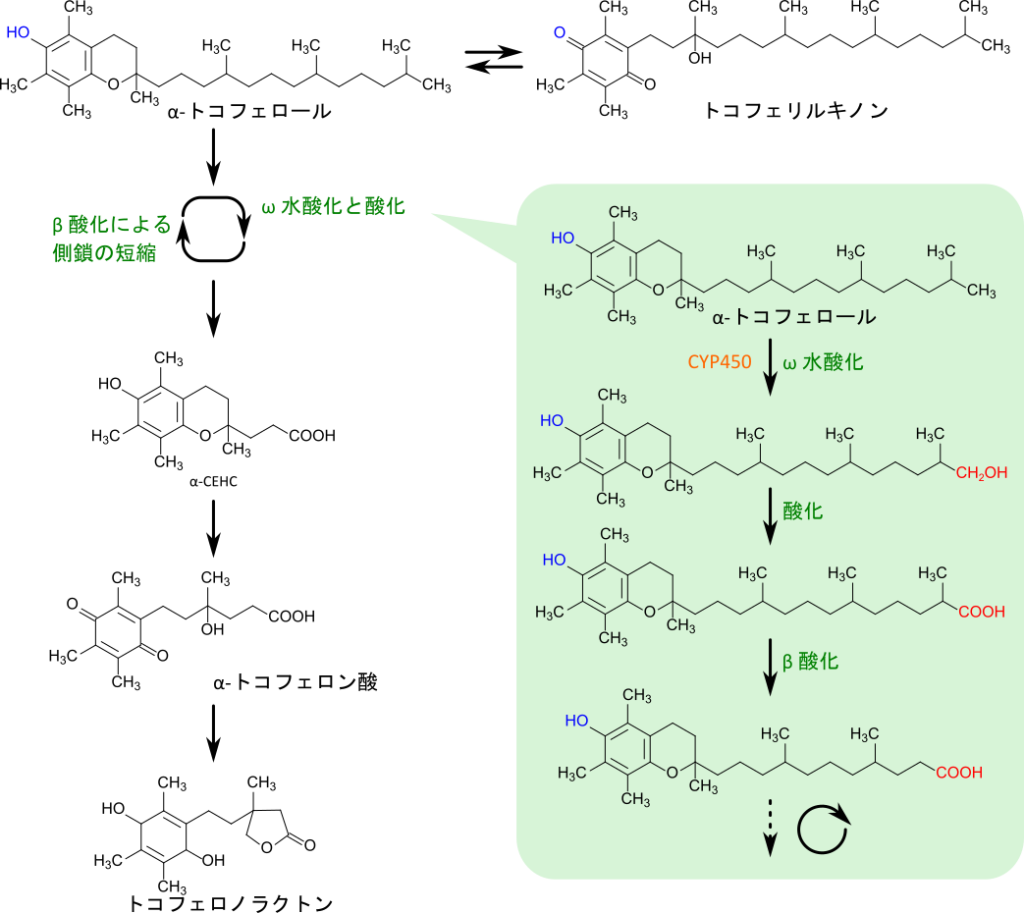
α-トコフェロールなどのビタミンEはω水酸化とβ酸化により炭素鎖が短縮され、トコフェロノラクトンやカルボキシエチルヒドロキシクロマン(CEHC)に代謝されると考えられています。出発物質がα体か、β体、γ体に対応して代謝産物はそれぞれ、α-CEHC、β-CEHC、γ-CEHCとなります。CEHC、特にγ-CEHCは単なるビタミンEの代謝産物というだけでなく、抗酸化作用や抗炎症作用、ナトリウム排泄の誘導などさまざまな生理活性を持つ分子でもあります(図 5)。

ビタミンEの異化経路
α-トコフェロールの異化経路は前述の通り、ω水酸化とβ酸化の繰り返しによると、考えられています。この過程では、トコフェロールの末端が水酸化された後、酸化を受けてカルボキシル化されます。その後、脂肪酸のようにβ酸化を受けて2炭素単位ずつ短くなっていくと考えられています。最終的にα-CEHCとなり、排泄されていきます。なお、α-CEHC が酸化されるとα-トコフェロン酸となりますが、水酸基がカルボキシル基を攻撃して環化することでトコフェロノラクトンへ変換され、尿中へ排泄されます(図 6)。
ビタミンC
ビタミンCの構造と特徴
ビタミンCは化学名をアスコルビン酸といい、ラクトン構造を持っています(図 7)。アスコルビン酸は容易に酸化されてデヒドロ-L-アスコルビン酸に変換されます(図 8)。このため、アスコルビン酸は活性酸素などの酸化剤を還元して、酸化作用から体内の細胞を守る役割を担っています。
また、水溶性であることから過剰摂取による弊害が起こりにくく、化学的に合成も可能であることから食品の酸化防止剤としてもよく利用されている物質です。

ビタミンC酸化物の異化と排泄
ビタミンC(アスコルビン酸)の異化経路
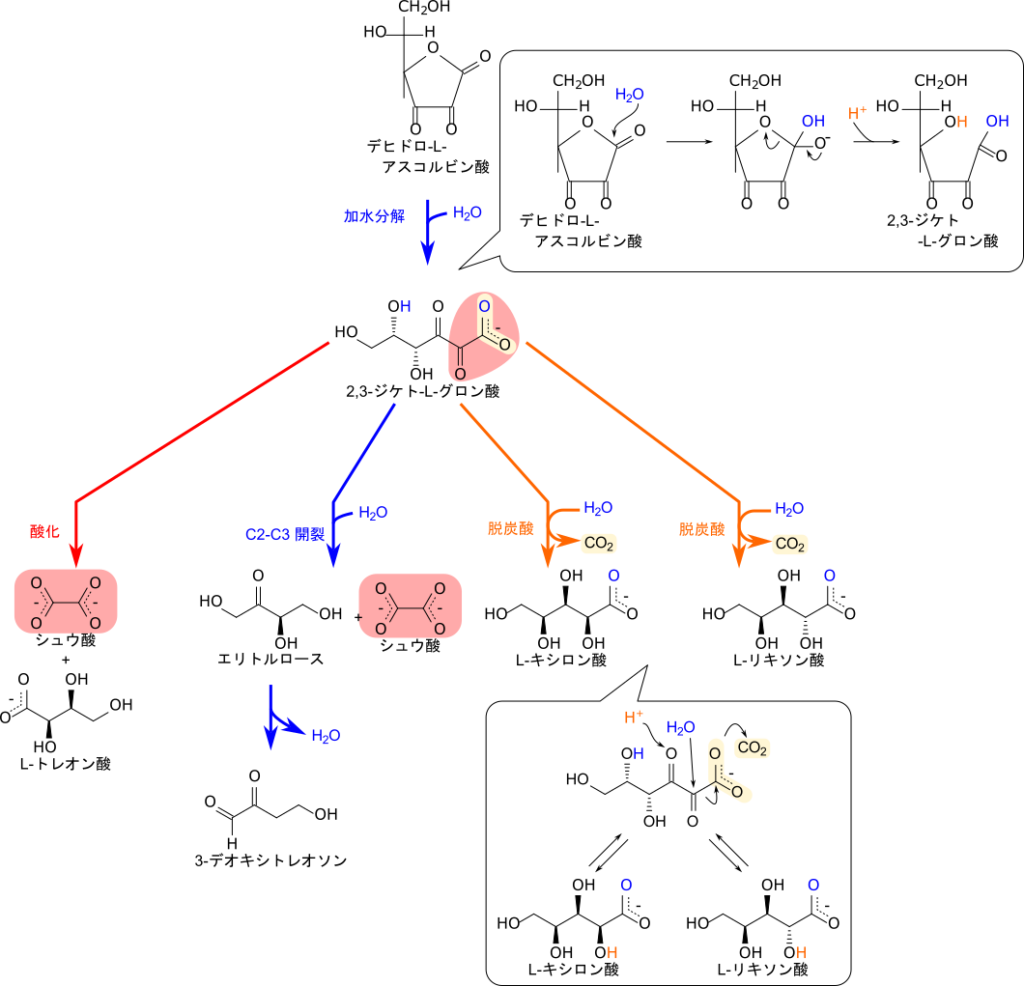
ラジカル種などを還元したアスコルビン酸はデヒドロ-L-アスコルビン酸に酸化されてしまいます(図 8)。デヒドロ-L-アスコルビン酸が加水分解を受けて開環すると2,3‐ジケト-L-グロン酸を生成します。この反応では、H2Oの酸素原子が1位の炭素を求核的に攻撃して結合し、続くプロトン移動によってラクトン環が開裂します。この2,3-ジケト-L-グロン酸は各種代謝産物への分岐点となっています(図 9)。
2,3-ジケト-L-グロン酸はさらに分解されることでL-リキソン酸やL-キシロン酸が生成されます。脱炭酸と水の付加反応を含む一連の反応によって1位のカルボキシル基が脱離して2位の炭素が新たなカルボキシル基となって、3位のカルボニル基周辺でプロトン移動が起こることでL-リキソン酸あるいはL-キシロン酸が生成されます。このとき、新たに生成される2位炭素上の水酸基の立体配置がどちらに向くかによって生成する分子が L-リキソン酸(図において紙面手前側に水酸基が来る場合)になるか L-キシロン酸(図において紙面奥側に水酸基が来る場合)になるかが決まります(図 9)。
一方で、2,3-ジケト-L-グロン酸が酸化などの反応によりC2-C3間で開裂するとトレオン酸とシュウ酸が生成されます。これらの代謝産物は最終的に尿中に排泄されます。C2-C3間の開裂では、エリトルロースとシュウ酸が生成される経路も存在します。エリトルロースはさらに酸化および脱水反応などを経て 3-デオキシトレオソンに変換されます(図 9)。
ビタミンC(アスコルビン酸)の異化産物とAGEs
アスコルビン酸の異化産物である 3-デオキシトレオソンは近年AGEs(advanced glycation end products)生成に関与していることが示唆されています。AGEs は非酵素的に糖化されたタンパク質や脂質などの総称であり、冠動脈硬化、心血管疾患、慢性腎疾患など健康上の問題との関連が指摘されている物質群です。アスコルビン酸の異化経路は研究途上にあり、ここに示した分子以外にも異化経路が提唱されています。これらの異化産物の中には3-デオキシトレオソンのようにAGEs 生成に関与するものも示唆されています。このような側面からもアスコルビン酸の異化産物は関心が集まっています。
この記事ではヒトでの異化経路を中心に解説しています。アスコルビン酸の異化経路は現在研究が進められている領域でもあり、様々な異化経路が提唱されています。また、植物と動物でも異化経路が異なっています。
アスコルビン酸とコラーゲン生成
コラーゲンは皮膚、血管、骨などを構造的に支える重要なタンパク質です。コラーゲンはプロリンが水酸化されて4-ヒドロキシプロリンになることで水を介してコラーゲンの3重らせん構造を安定化することができます。この重要な水酸化は翻訳後修飾でプロリルヒドロキシラーゼによって行われます(図 10)。
プロリン残基の水酸化機構
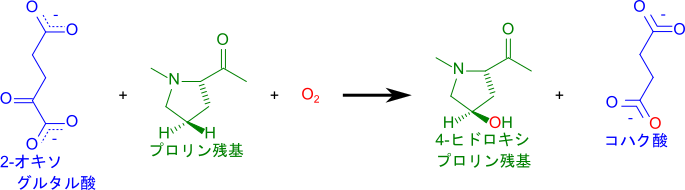
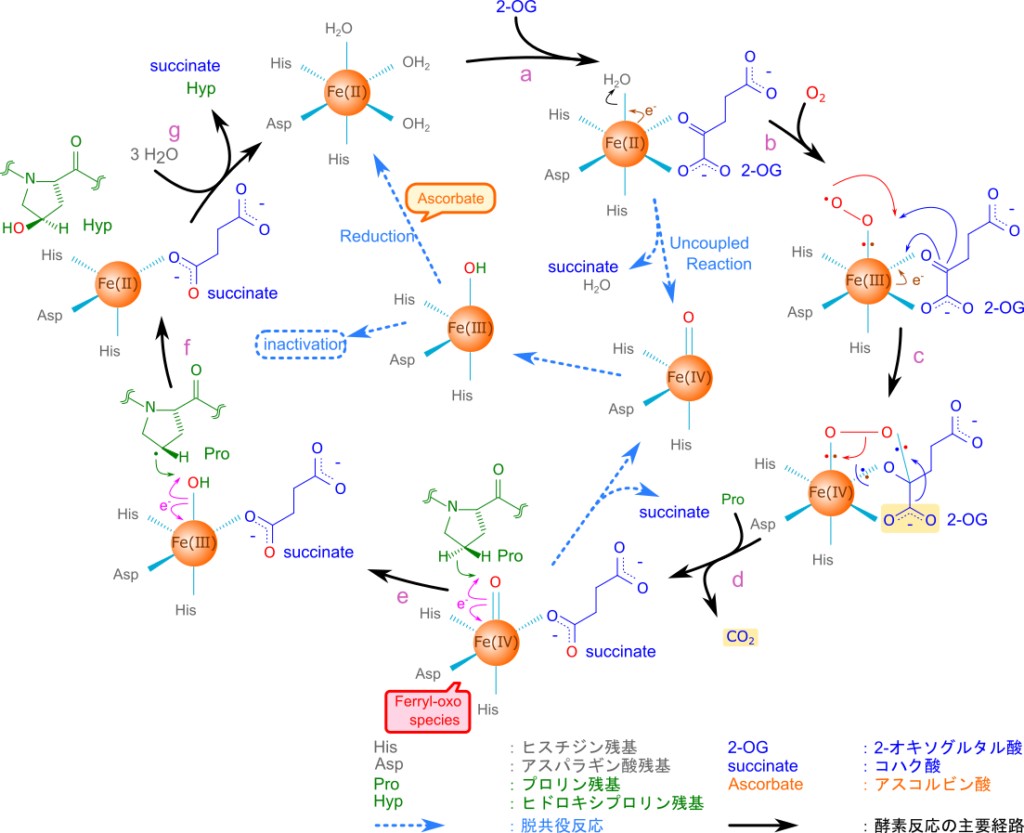
プロリルヒドロキシラーゼ(prolyl hydroxylase)はFe(Ⅱ) を分子内に持っており、この鉄原子が反応において中心的な役割を担います。鉄原子はヒスチジン、アスパラギン酸、水が配位して、八面体配位構造をとっています。基質である2-オキソグルタル酸と酸素が供給されると、2-オキソグルタル酸の酸化的脱炭酸(Oxidative decarboxylation)とFe 中心における O2 活性化が共役して反応が進行し、2-オキソグルタル酸は CO2 とコハク酸に変換されます。この時、酸素分子の一方の酸素原子はコハク酸に取り込まれ、もう一方は鉄原子と鉄-オキソ種(ferryl-oxo species)を形成します(図 11、a~d)。この時鉄原子は電子を供与して Fe(Ⅳ) に酸化されます。生成された鉄-オキソ種は強い酸化剤として機能し、プロリンの水素原子を引き抜き(Hydrogen atom transfer; HAT)、水酸基の再結合により水酸基をプロリンに導入してヒドロキシプロリン残基を生成します。この過程では一度 Fe(Ⅳ) となった鉄原子が一連の反応の結果、電子を受け取って Fe(Ⅱ) へ還元されます。最後にコハク酸が放出されて水分子が配位し、初期状態に戻ります(図 11、d~g)。このように鉄原子はプロリンの水酸化において電子のやり取りに深く関与しています。
プロリルヒドロキシラーゼ活性に対するビタミンCの役割
この反応では理論上、Fe(Ⅱ) に戻りますが、基質が不足すると脱共役反応(uncoupled reaction)が起こってFe(Ⅲ) が生成される場合があります。この場合、プロリルヒドロキシラーゼは最終的に不活性化されてしまいます(図11、青矢印)。アスコルビン酸は Fe(Ⅲ) を Fe(Ⅱ) に還元することでプロリルヒドロキシラーゼの活性を維持する役割を担っています。アスコルビン酸は脱共役反応による酵素の不活性化を防ぎ、コラーゲンの翻訳後修飾が速やかに進むことに貢献しています(図11)。
ビタミンEとビタミンCの協調的な機能
ビタミンEは細胞膜やLDLなどのリポタンパク質において脂質ラジカルを還元し、脂質酸化の進行を抑制する役割を担うことは上述の通りです。一方でこの過程で生成されるビタミンEラジカルは脂質ラジカルを生成する可能性もあるため、速やかに還元される必要があります。ビタミンCはビタミンEラジカルを還元してビタミンEに変換することにより、脂質ラジカルの産生を抑制するだけでなく、ビタミンEを再生する役割を担っています。実際に、ビタミンCとビタミンEを同時に作用させることにより脂質酸化を効率的に抑制できることが in vitro、in vivo ともに実験的に示されています。このようにビタミンCとビタミンEは協調的に働くことにより細胞膜やリポタンパク質内の脂質酸化を効率的に抑制しています(図 12)。
練習問題
アスコルビン酸の酸化産物を以下の中から選べ
プロリルヒドロキシラーゼによるプロリンの水酸化について正しい記述を選べ
ビタミンEとビタミンCが協調的に機能することの意義として正しい記述を選べ
ビタミンCとビタミンEは、単に「抗酸化ビタミン」として並べて語られるだけの存在ではありません。脂質膜でラジカル連鎖を食い止めるビタミンE、水溶性環境でラジカルを処理し、さらにビタミンEを再生するビタミンC――両者はそれぞれ異なる場所で役割を分担しながら、協調的な抗酸化作用を実現しています。
さらに、これらのビタミンは単に酸化を防ぐだけでなく、異化産物の生成、コラーゲン形成、さらにはAGEs生成との関わりなど、多様な生命現象とも深く結びついています。特にビタミンCの異化経路やビタミンE代謝産物の機能は、現在も研究が進められている興味深い領域です。
普段「ビタミン」とひとまとめにされがちなこれらの分子も、その働きを分子レベルで追っていくと、脂質膜、ラジカル、鉄イオン、酵素反応、リポタンパク質輸送など、多くの分子が連携する精巧なシステムの一部であることが見えてきます。
この記事を通して、ビタミンCとビタミンEを「栄養素」としてだけではなく、生体内で連携して働く動的な分子ネットワークとして捉えるきっかけになれば幸いです。
参考文献
共通
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp. 391-415, 751-771
- D. Voet, J. G. Voet, C. W. Pratt 著、田宮信雄、八木達彦、遠藤斗志也、吉久徹訳 (2017) ボート 基礎生化学 第5版. 東京化学同人. pp. 84-117, 294, 164-195, 440-476
- Peter J. Kennelly, Kathleen M. Botham, Owen P. McGuinness, Victor W. Rodwell, P. Anthony Weil 著、清水孝雄、水島昇 監訳 (2024). イラストレイテッド ハーパー・生化学 原書32版. 丸善出版. pp. 633-650
- 五十嵐脩、江指隆年 編 (2011). ビタミン・ミネラルの科学. 朝倉書店. pp. 1–39
ビタミンE
- Ogawa, S., & Iuchi, K. (2024). α-Tocopherol: New Perspectives and Challenges for Achieving the Sustainable Development Goals (SDG) Target. In Journal of Oleo Science (Vol. 73, Issue 4, pp. 519–538). Japan Oil Chemists Society. https://doi.org/10.5650/jos.ess23199
- Wu, J. H., & Croft, K. D. (2007). Vitamin E metabolism. Molecular Aspects of Medicine, 28(5–6), 437–452. https://doi.org/10.1016/j.mam.2006.12.007
- YAMAUCHI, R. (1997). Vitamin E: Mechanism of Its Antioxidant Activity. Food Science and Technology International, Tokyo, 3(4), 301–309. https://doi.org/10.3136/fsti9596t9798.3.301
ビタミンC
- Kuiper, C., & Vissers, M. C. M. (2014). Ascorbate as a cofactor for Fe-and 2-oxoglutarate dependent dioxygenases: Physiological activity in tumour growth and progression. Frontiers in Oncology, 4(NOV), 117223. https://doi.org/10.3389/fonc.2014.00359
- Dewhirst, R. A., & Fry, S. C. (2018). The oxidation of dehydroascorbic acid and 2,3-diketogulonate by distinct reactive oxygen species. Biochemical Journal, 475(21), 3451. https://doi.org/10.1042/BCJ20180688
- Osipyants, A. I., Poloznikov, A. A., Smirnova, N. A., Hushpulian, D. M., Khristichenko, A. Y., Chubar, T. A., Zakhariants, A. A., Ahuja, M., Gaisina, I. N., Thomas, B., Brown, A. M., Gazaryan, I. G., & Tishkov, V. I. (2018). L-ascorbic acid: A true substrate for HIF prolyl hydroxylase? Biochimie, 147, 46–54. https://doi.org/10.1016/j.biochi.2017.12.011
- You, Z., Omura, S., Ikeda, H., Cane, D. E., & Jogl, G. (2007). Crystal Structure of the Non-heme Iron Dioxygenase PtlH in Pentalenolactone Biosynthesis. Journal of Biological Chemistry, 282(50), 36552–36560. https://doi.org/10.1074/jbc.M706358200
- Linster, C. L., & van Schaftingen, E. (2007). Vitamin C: Biosynthesis, recycling and degradation in mammals. In FEBS Journal (Vol. 274, Issue 1, pp. 1–22). https://doi.org/10.1111/j.1742-4658.2006.05607.x
- Nemet, I., & Monnier, V. M. (2011). Vitamin C degradation products and pathways in the human lens. Journal of Biological Chemistry, 286(43), 37128–37136. https://doi.org/10.1074/jbc.M111.245100
- Smirnoff, N. (2018). Ascorbic acid metabolism and functions: A comparison of plants and mammals. In Free Radical Biology and Medicine (Vol. 122, pp. 116–129). Elsevier Inc. https://doi.org/10.1016/j.freeradbiomed.2018.03.033