多くのアミノ酸と異なり、ヒスチジンの生合成経路はプリン塩基やピリミジン塩基の合成と深く関連しています。この事実は、この経路に初期生命の代謝進化の痕跡が残っていることを示唆しています。本記事では、ヒスチジンの生合成経路を解説するとともに、原始生命に関するRNA ワールド仮説とこの経路の関連についても紹介します。
ヒスチジンの生合成経路
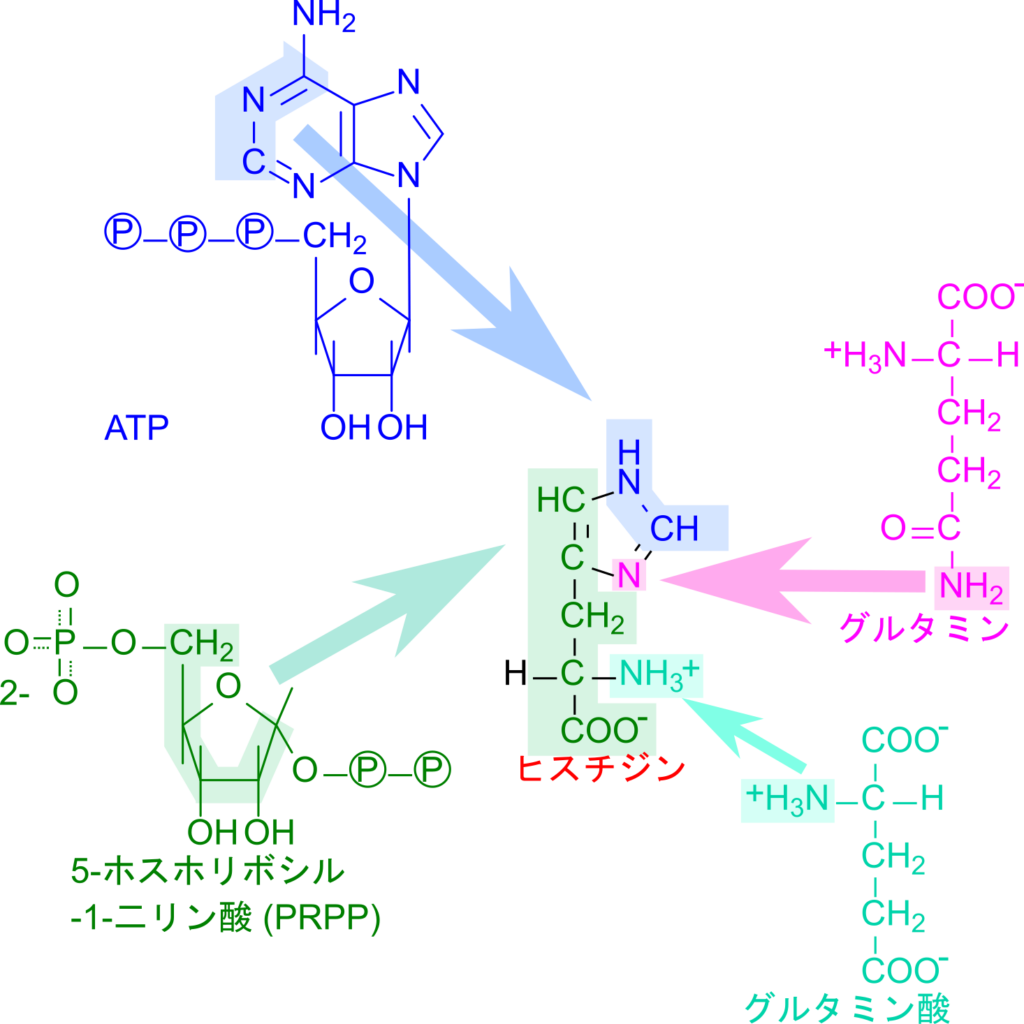
ヒスチジンの生合成経路は少し複雑なので、ヒスチジンを構成する原子の由来を確認していきましょう。ヒスチジンを構成する原子は二つの窒素原子を除いて ATP と 5-ホスホリボシル-1-二リン酸(PRPP)に由来しています。主鎖から側鎖の一部は PRPP から供給され、側鎖のイミダゾール環の一部は ATP から供給されます。イミダゾール間の窒素原子はグルタミンの側鎖の窒素原子に由来し、α 炭素に結合しているアミノ基はグルタミン酸からアミノ基転位を受けて供給されます(図1)。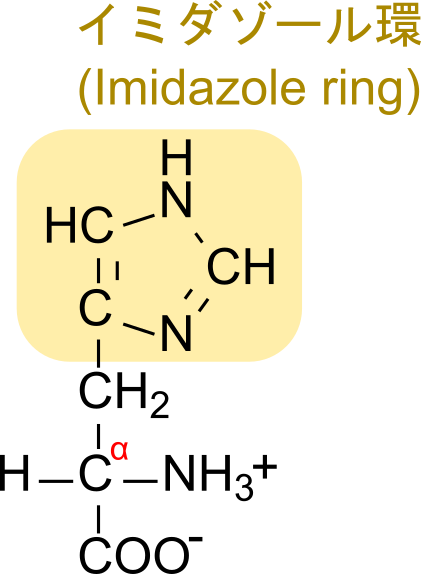
このことから、ヒスチジンの生合成は PRPP と ATP が会合したのちに、不必要な部分が切除され、アミノ基が各分子から転位されるという意外とシンプルな流れで行われています。それでは詳細についてみていきましょう。
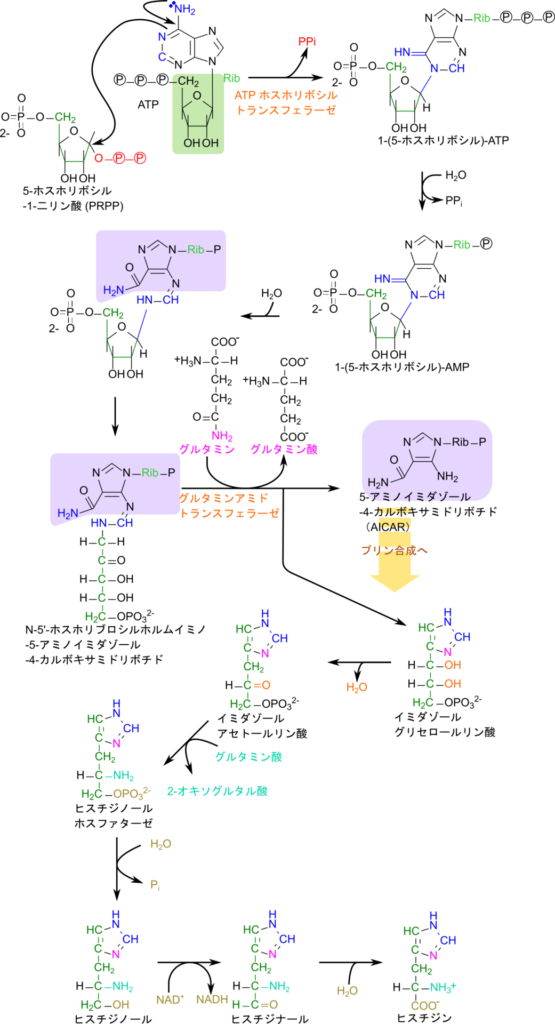
ヒスチジンの生合成経路は PRPP と ATP が会合することから始まります。この会合反応はプリン環の窒素原子が PRPP の炭素を求核的に攻撃して結合を形成し、アミノ基の非共有電子対が遷移してイミノ基に変換されることで進みます。その後、ATP 部分の三リン酸の内、二リン酸が遊離すると 1-(5-ホスホリボシル)-AMP が生成されます。この反応では遊離したピロリン酸が即座にリン酸基まで分解されることで、ATP 二分子分のエネルギーを産生します。このエネルギー分、反応の前後で化学ポテンシャルの差が大きくなるため、この反応は不可逆的となります。その後、プリン環の六員環部分が開環したのちに、リボース部分が開環すると N-5′-ホスホリブロシルホルムイミノ-5-アミノイミダゾール-4-カルボキサミドリボチドが生成されます。この物質は、5-アミノイミダゾール-4-カルボキサミドリボチド部分とイミダゾールグリセロールリン酸の前駆体となる部分から構成されています。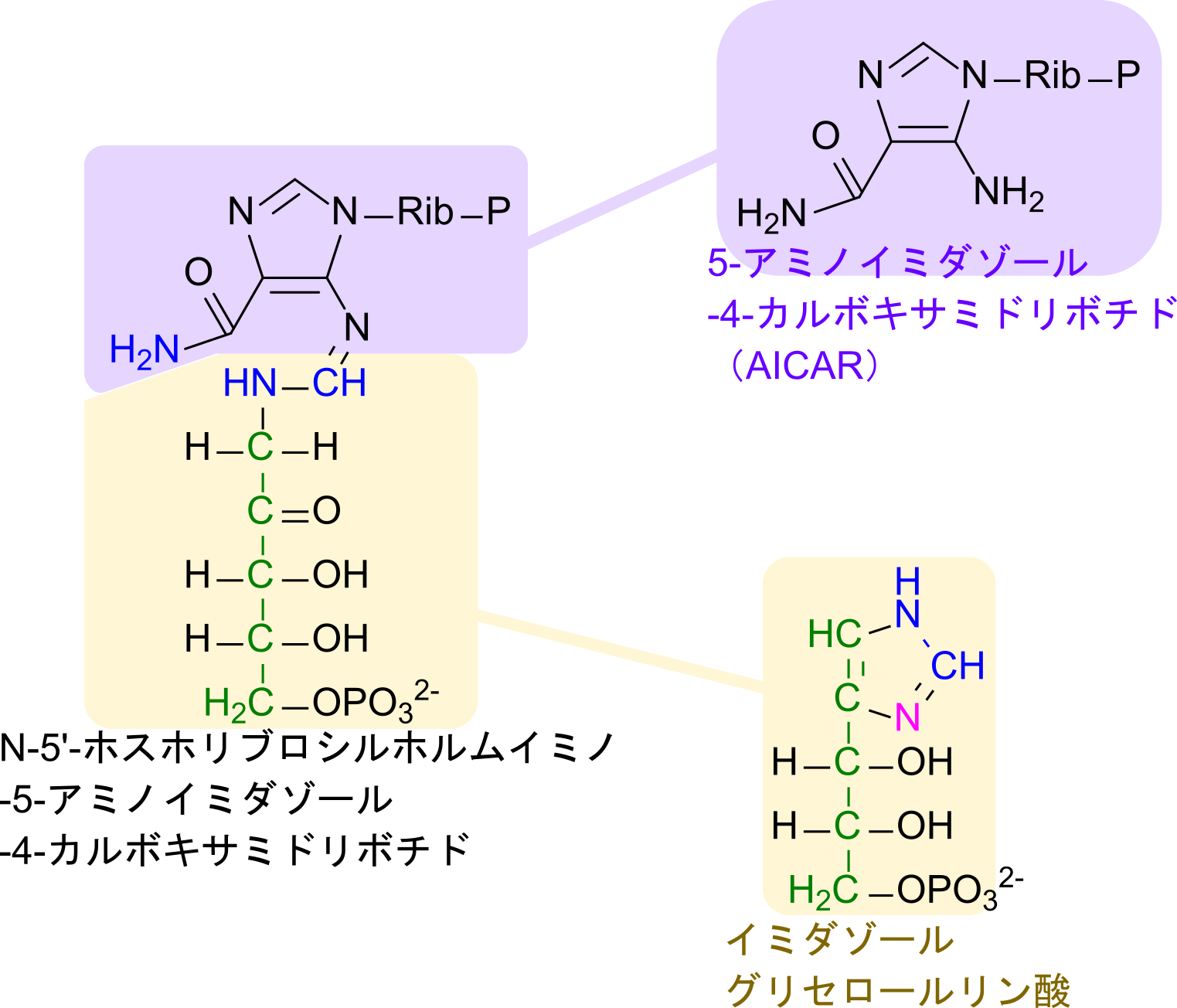
そこで、グルタミンからのアミノ基転位を受けるとともに分解を受けて、5-アミノイミダゾール-4-カルボキサミドリボチド(AICAR)とイミダゾールグリセロールリン酸に分解されます。ここで、産生される AICAR はプリン塩基の中間体でもあります。一方で、イミダゾールグリセロールリン酸から、水が遊離するとイミダゾールアセトールリン酸に変換されます。ここで、カルボニル基がグルタミンに由来するアミノ基転位のターゲットとなります。アミノ基が転位されたのち、リン酸が遊離するとヒスチジノールとなります。最後に末端のアルコール部分を酸化するとヒスチジンが生成されます(図2)。
ヒスチジンの特殊性とRNA ワールド仮説
ヒスチジンは PRPP から合成されますが、PRPP はプリン塩基やピリミジン塩基の生合成にも利用されています(詳細はこちら)。また、ATP もヒスチジンの合成に関与しており、さらには、ヒスチジンの生合成過程で生じる AICAR はプリン塩基生合成の中間体でもあります。このようにヒスチジンは核酸(特に RNA)と関連の深い分子です。一方で、ヒスチジンは窒素原子を含むことから求核基として機能することやプロトンの供給と受容が可能であることなどから、多く酵素の活性部位に利用されるアミノ酸です。また、プリン塩基の五員環はイミダゾール環となっていますが、RNA 酵素ではこのイミダゾール部分がヒスチジンと同様な働きを持つと考えられます(ちなみに、イミダゾール環はヒスチジンの側鎖でもありますのでここからもヒスチジンと RNA の類似性がうかがえます)。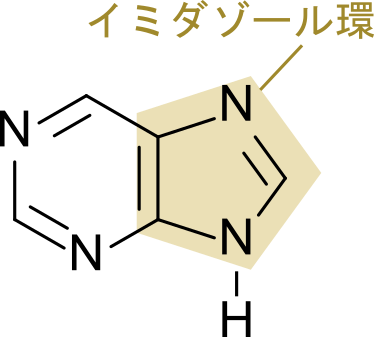
これらの事実はかつて生命が RNA を基礎に誕生したという仮説を支持していると考えられています。
かつての生命は RNA を中心としていたという仮説があります。この仮説の要点は以下の二点です。
- DNA による情報の保存はもともと RNA が担っていた
- タンパク質による触媒能はもともと RNA が担っていた
このような仮説は RNA ワールド仮説(RNA World Hypothesis)と呼ばれます。よく知られているように、RNA は情報を保存する能力があります。一方で、RNA には化学反応を触媒する能力も有しています。特に近年ではリボザイム(ribozyme)と呼ばれる触媒能を持つ RNA が発見されており、RNA にも化学反応を触媒する能力があることが証明されています。例えば、リボソーム RNAはタンパク質合成においてペプチド結合を形成する反応を触媒しています。また、ハンマーヘッドリボザイムはウイルスや住血吸虫の RNA の複製に関与しています。
ところが、RNA は構造的な問題から切断を受けやすく不安定であるという弱点があります(くわしくはこちら)。この問題は情報を保存する物質としては致命的です。このため、長い進化の中で、情報の保存をより安定なDNAに渡していったと考えられます。触媒能についても同様で、RNA では配列を構成するヌクレオチドがたった四種類しかないのに対してタンパク質の場合は 20 種類もあり、その中にはヒドロキシ基(-OH 基)、メルカプト基(-SH 基)、アミド基(-NH2 基)、カルボキシ基(-COOH 基)など様々な置換基を持つものがあります。生体内で行われる幅広い反応をカバーするためにはバリエーションに優れたタンパク質の方が有利です。その結果、RNA の持つ触媒能はタンパク質に譲っていきました。ヒスチジンの生合成経路が核酸と深く関連していることは RNA からタンパク質に触媒能が移っていった名残とみることができます。このように、ヒスチジンが持つ特殊性はかつて RNA が生命の中核を担っていたことを示す証拠の一つと解釈することができるのです。
5-アミノイミダゾール-4-カルボキサミドリボチドの名称について
5-アミノイミダゾール-4-カルボキサミドリボチドについて命名を確認しておきます。この物質はプリン塩基に利用される重要な分子ですが、名前が少し複雑なので確認していきましょう。
まず、語尾がリボチドとなっています。リボチドとはリボヌクレオチドのことを言っています。リボヌクレオチドとは糖部分がリボースであり、リン酸基が結合していて塩基が結合している構造を言います。今回の場合、塩基部分はプリン塩基やピリミジン塩基でありませんが、五員環とカルボキサミド、アミノ基で構成される部分(青枠部分)はアミノ基を有する環状構造を持つことから塩基とみなせます。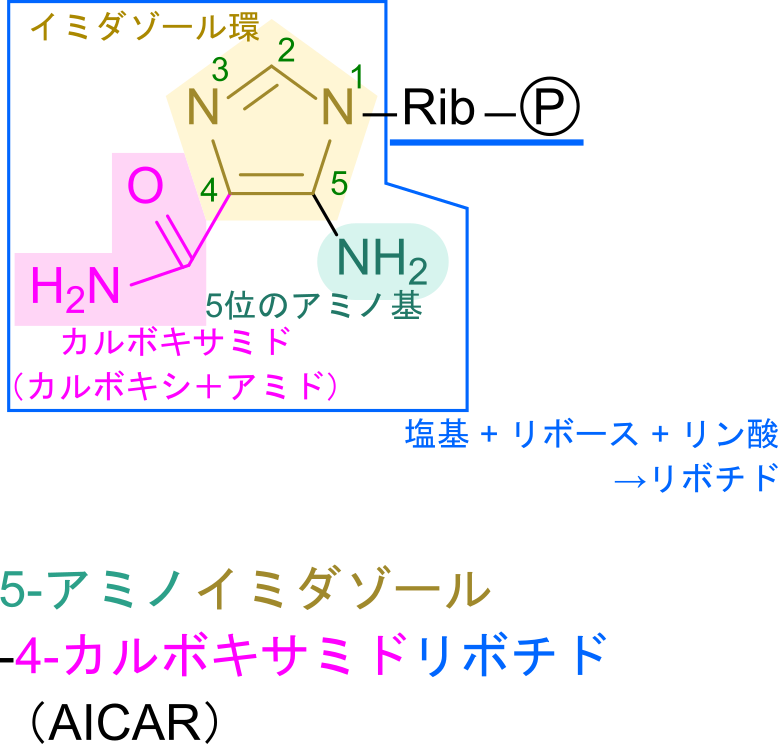
従って、5-アミノイミダゾール-4-カルボキサミドリボチドは全体としてリボヌクレオチドの構造をしているため、”リボチド”という語が末端についています。次に、五員環部分はイミダゾール環と呼ばれる構造です。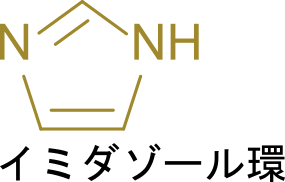
イミダゾール環はヒスチジンの五員環部分でもあります。番号は -NH 基を1として反時計回りにつけていきます。なお、5-アミノイミダゾール-4-カルボキサミドリボチドではリボースが結合している窒素が -NH 基に相当します(上の構造と見比べてみてください)。次に 5-アミノイミダゾールについてみていきます。この意味は 5 位の炭素にアミノ基(-NH2)がついていることを意味します。次に 4-カルボキサミドについて考えます。カルボキサミドとはカルボキシ+アミドであり、CO-NH2 の構造を指します。このカルボキサミドが 4 位の炭素に結合していることから、4-カルボキサミドとなっています。以上の内容を合わせて命名を考えると 5-アミノイミダゾール-4-カルボキサミドリボチドとなります。なお、英語名では 5-AminoImidazole-4-Carboxamide Ribotide となり、AICAR と略されます。
ヒスチジンの生合成経路は、他のアミノ酸とは一線を画す独自性を備えています。解糖系や TCA 回路に由来する多くのアミノ酸と異なり、ヒスチジンは PRPP や ATP といった核酸と深く関わる分子から合成され、その過程で AICAR のようなプリン塩基の中間体が生じます。こうした事実は、ヒスチジンが RNA と極めて密接な関係をもつアミノ酸であることを示しています。
さらに、この合成経路は、RNA がかつて生命活動の中心であったとされる「RNAワールド仮説」を支持するものであり、ヒスチジンはその名残をとどめる重要な手がかりとも考えられています。
本記事では、ヒスチジンの構造や生合成経路をわかりやすく解説するとともに、その背後にある生命進化の痕跡にも目を向けました。分子の構造から過去の生命像を読み解くという、少し変わった視点からヒスチジンを見つめることで、代謝経路が単なる反応の連なりではなく、生命の歴史を物語る重要な証拠であることに気づいていただけたのではないでしょうか。
参考文献
- 島原健三 (1991). 概説 生物化学. 三共出版. pp. 208-223
- Jeremy M. Berg, John L. Tymoczko, Gregory J. Gatto Jr., Lubert Stryer著、入村達郎、岡山博人、清水孝雄、中野徹訳 (2018). ストライヤー生化学 第8版. 東京化学同人. pp.665-693
- D. Voet, J. G. Voet, C. W. Pratt 著、田宮信雄、八木達彦、遠藤斗志也、吉久徹訳 (2017) ボート 基礎生化学 第5版. 東京化学同人. pp.496-503
- KEGG PATHWAY DATABASE. 2025-03-03. https://www.genome.jp/kegg/pathway.html
- Robertson, M. P., & Joyce, G. F. (2012). The Origins of the RNA World. Cold Spring Harbor Perspectives in Biology, 4(5), a003608–a003608. https://doi.org/10.1101/cshperspect.a003608